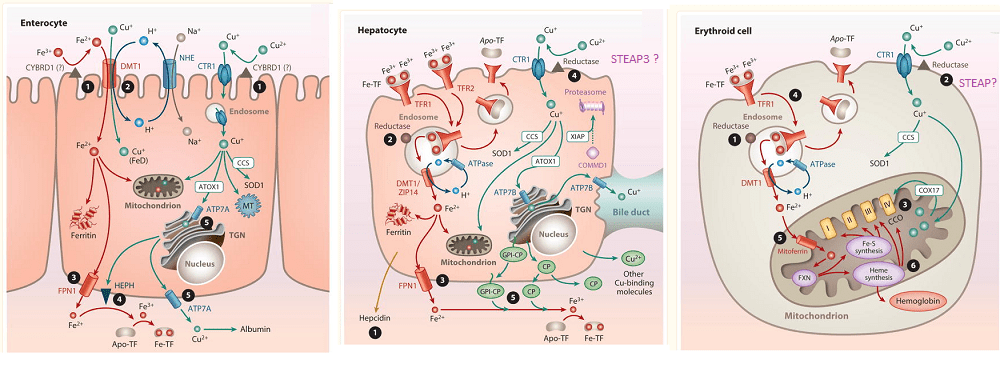
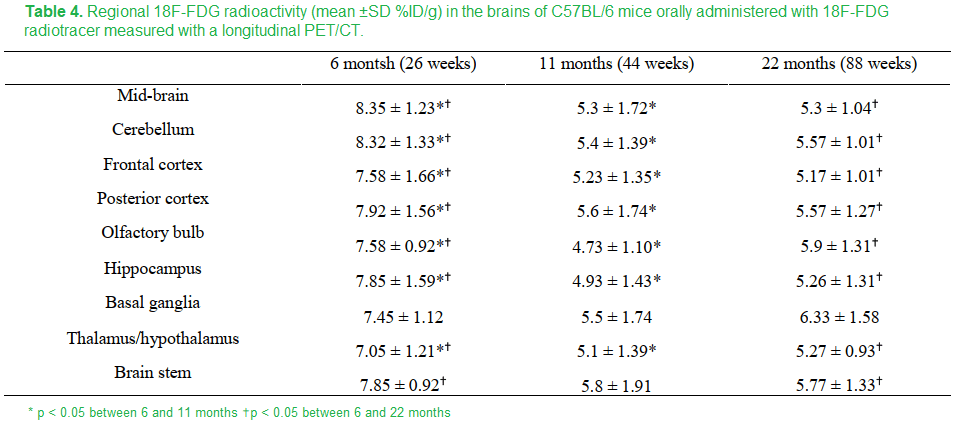This post addresses the existence of a putative Cu(II) reductase that sits adjacent to the Ctr1 Cu+ channel. Sukru Gulec and James Collins wrote an excellent review on absorption of iron (Fe) and copper (Cu). This is a composite of three images going from the entry of Cu in the duodenum, transfer to the hepatocyte, and the red blood cell, erythroid cell.
Images from Gulec& Collins (2014), Liberty has been taken to write the names of putative Cu(II) from the figure legends of these images from the public access manuscript.
Let us start our journey with the
CYBDR1 , aka DCytB is a small membrane protein for which UniProt lists the following catalytic function:
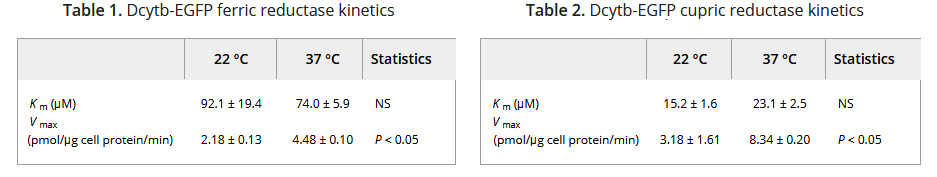
Note that Cybdr1 has a lot in common with cytochrome P450s in the liver, hence the “Cy” as part of the name. The “D” must have come from “duodenum,” a location in the proximal small intestine responsible for absorbing Fe and Cu. The 31 kDa Cybdr1 gene was fused to the gene for Green Fluorescence protein and expressed in a kidney cell line. The kinetics of reducing Fe3+ and Cu2+ was measured. [2] The Fe(II)-ferrozine complex formation was detected by the absorbance at 562 nm. Capture of Cu(I) by BCS was monitored by the change in absorbance at 482 nm. [2]
Data from ref [2]
This particular kidney cell line expresses the divalent metal ion tansporter, DMT1. The authors demonstrated that the Kcyb1 transfected cells had absorbed 5x as much 59Fe originally added in the +3 ferric oxidation state as non transfected cells. [2] The authors cited literature showing that DCYTB knockout mice seemed normal and that no mutations in the human DCYTB gene have been associated with human disease at the time of the manuscript published in 2008. [2]
STEAP: Six Transmembrane Epithelial Antigen of Prostate
In 2006 a group from Millennium Pharmaceuticals and Children’s Hospital of Harvard characterized the tissue expression and reductase activity of Steap isoforms. [3] Tissue expression of STEAP1-4 was determined by use of radioactive RNA hybridization probes and whole body sections of mice. STEAP1-4 were expressed in HEK293 cells. [3] Fe3+ reduction to Fe2+ was measured with ferrozine, that absorbs 562 nm. The reduction of Cu2+ to Cu+ was measured with BCA that absorbs at 482 nm . [3]
From ref [3] with minor modifications for easier reading.
Note that Cu2+ uptake was already pretty high with just the empty vector used to insert the Steap gene into HEK293 cells. Cu2+ does seem to compete with Fe3+ for reductase activity.
Uptake of both metal ions was measured with radioactive analogs. [3]
the final figure from [3] with some cell line specific mRNA expression data from Protein Atlas. HepG2 cells are a liver cell line. Caco-2 are an intestinal line.
All in all, these data are really not that convincing that the Steap and DCyb1 reductases are really doing that much for copper absorption.
References
Gulec, S., & Collins, J. F. (2014). Molecular mediators governing iron-copper interactions. Annual review of nutrition, 34, 95–116. PMC free article
Wyman S, Simpson RJ, McKie AT, Sharp PA. Dcytb (Cybrd1) functions as both a ferric and a cupric reductase in vitro. FEBS Lett. 2008 Jun 11;582(13):1901-6. Free article
Ohgami R. S., Campagna D. R., McDonald A., Fleming M. D. (2006) The Steap proteins are metalloreductases. Blood 108, 1388–1394 PMC free article
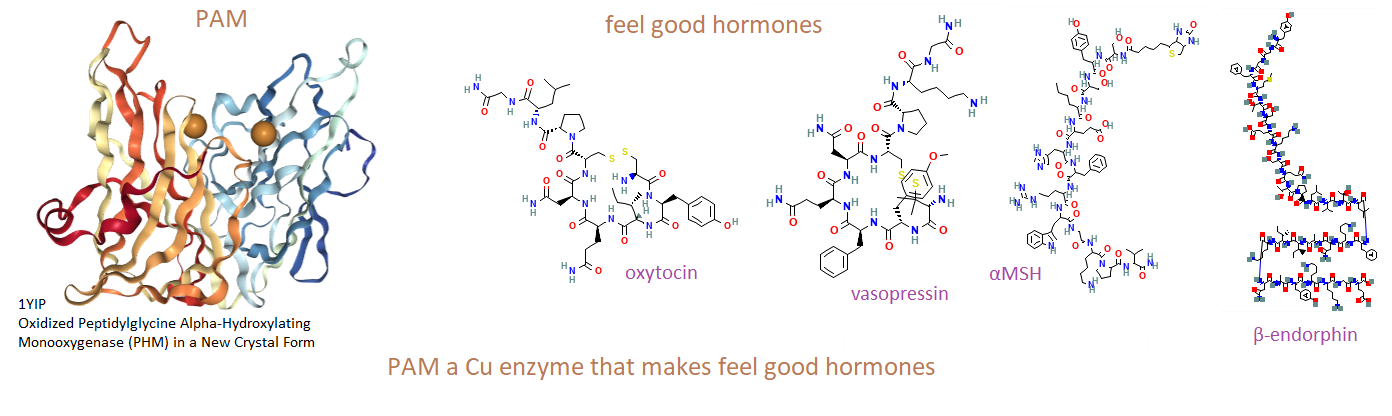
This post is about peptide hormones that make us feel good. It is not about dopamine and serotonin… The copper enzyme Peptidylglycine α-Amidating Monooxygenase (PAM) is responsible for the clipping of peptides to form common hormones we know as vasopressin, oxytocin, α-MSH, and β-endorphin. [1]
How PAM works
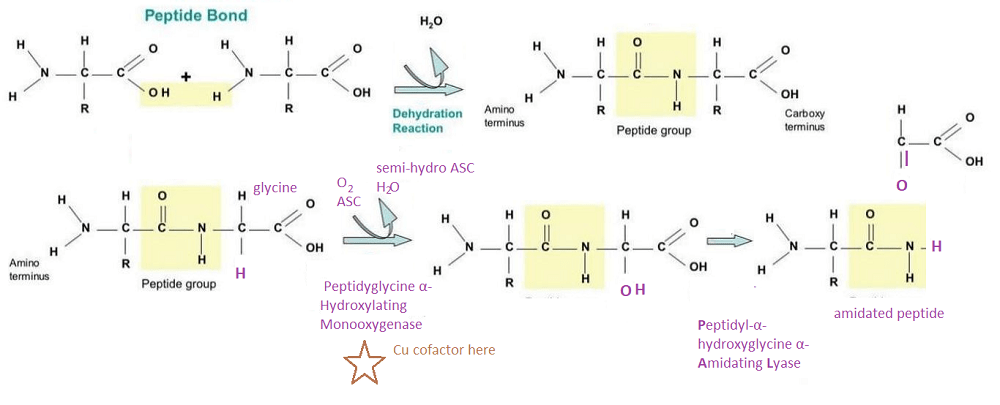
PAM is actually two enzymes in one. PHM uses copper to tag with oxygen. PAL snips at the tagged carbon.
Top, How a peptide bond is formed. R can be one of 20 groups unique to each amino acid. This includes H for glycine. Bottom: The enzyme PAM
The top panel covers what a peptide bond is. The bottom panel describes how the dual function enzyme PAM tags and clips the peptide resulting in the N-terminal (butt) of glycine. This is just something to think about. Copper is required for the production of some very important peptide hormones. We still do not know why the C-terminal amidation is so important. Many think it neutralizes the charge and increases binding to the hormone receptor.
PAM+/- mice are just messed up
A 2011 study found that mice with only one functional copy of the PAM gene, PAM+/-, had increased anxiety and decreased cold tolerance. [2] Copper deficiency in wildtype mice mimicked these symptoms. Discussion was given to PAM sometimes ending up in the nucleus. Gene transcripts for copper handling proteins were found to be altered in two key locations: the heart and the pituitary gland. Note that Thyrotropin-releasing hormone (TRH) is released from hypothalamic neurons that act on neurons of the pituitary gland.. that product ACTH, that acts on the adrenal gland to release cortisol… All of these feeds back on TRH.
highlights from reference [2] TRH-Gly has not been cleaved by PAM.
Why these mice felt anxious and whether the bad feelings were due to TRH-Gly getting stuck in the hypothalamus was not really a topic of discussion. Just that the hypothalamus connects to the pituitary gland brings us to the next topic.
PAM and amidated pituitary hormones
Some quick comments before getting into hard core science.
Oxytocin is the cuddling a cute puppy or a baby peptide. It is involved in all aspects of making more of one’s own species…. or a suitable surrogate of another species.
Vassopressin, also arginine vasopressin (AVP), is released by diurnal variations in sunline as well as changes in the osmolality (salt content) of the blood. AVP is similar to oxytocin, but does some very different things too such as increasing water channels in the kidney to aid in water absorption. The Wikipedia page on Vassopressin does a good job of comparing and contrasting AVP and oxytocin.
α-Melanocyte stimulating hormone, or α-MSH, is what tells our melanocytes to start producing melanin in response to sunshine. Is α-MSH the “Sunshine on my shoulders makes me happy” hormone? Maybe yes, maybe no. α-MSH is cleaved from same precursor as
Endorphinsbind to the same receptors as opioid pain killers. Endorpins are the eating chocolate, listening to music, meditation, sex, laughter, and sunshine hormones.
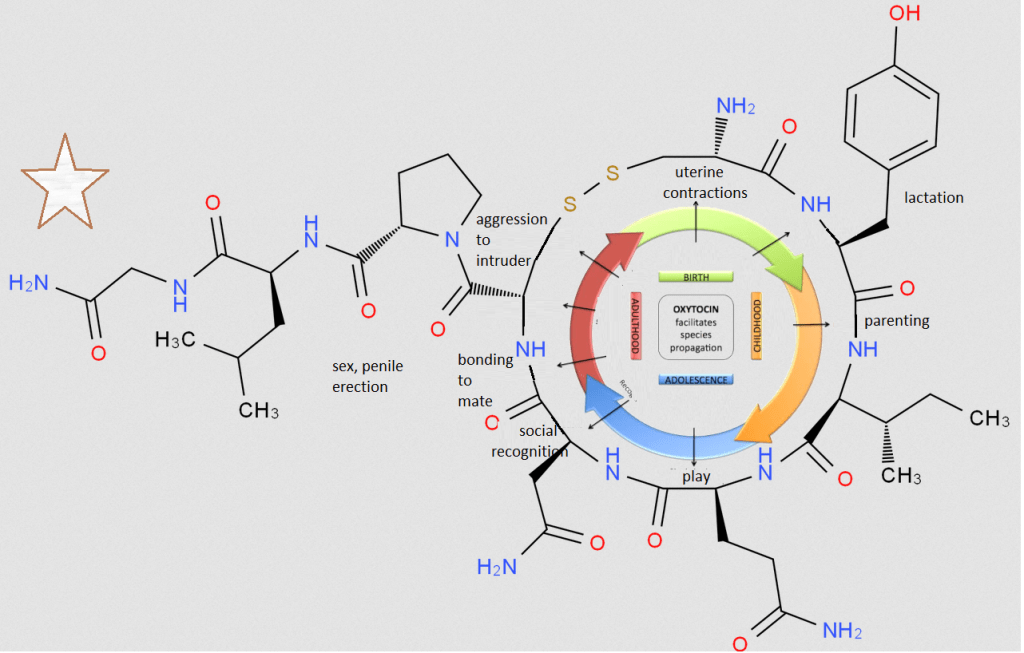
Oxytocin
A pictorial overview of oxytocin throughout the lifespan of the animal.
Adapted from Lee 2009 {3}
A study from 1992 demonstrated that guinea pig dams made copper deficient (0.8 microgram Cu/g diet) required 0.5 to 6.2 U oxytocin to injections to induce delivery of pups. [4] Many of the pups died of internal hemorrhaging, possibly due to lack of lysyl oxidase crosslinking of collagen. [4] In Cu adequate dams, pup survival was at 79%, Only 28% of pups survived in copper deficient dams.
In another study pregnant rats were placed on the copper-deficient or copper-adequate treatment 7 days after mating. The “purified” diet contained 0.35 mg Cu/kg and 42 mg Fe/kg by chemical analysis. Offspring and dams on the copper-deficient treatment drank deionized water, whereas copper adequate treatment groups drank water that contained 20 mg Cu/L by adding CuSO4 to the drinking water. [5] Decreased PAM activity showed up in the mid brains and atria of the heart. [5] We will not get into whether Cu(II) in drinking water is optimally bioavailable. Decreased superoxide dismutase SOD and cytochrome C oxidase CCO activity was noted.
Vasopressin aka AVP
The Yoshimura review made no mention of PAM being involved in AVP synthesis. They did state that AVP is made in the hypothalamus in response to stress. [5] The reader is invited to consult [6] for a comprehensive review of the tissue distribution of the three receptors of AVP and their physiological functioning. The Song and Albers (2018) review made the point that many rodent studies have demonstrated that oxytocin and AVP can bind to each others receptors, all of which are G-protein coupled receptors. [7] IF we can trust Wikipedia, AVPR1A couples with Gαq/11, AVPR1B couples with Gαq/11 , and AVPR2 with Gαs, while the oxytocin receptor couples with Gαq.
Three variety of alpha subunits that couple to heterotrimeric G protein coupled receptors. The other two units of the heterotrimer has another function.
In summary, AVP and oxytocin are very similar in sequence and structure. They can bind to the same receptors. The oxytocin and AVPR1 rceptors have similar down stream signalling via PLC.
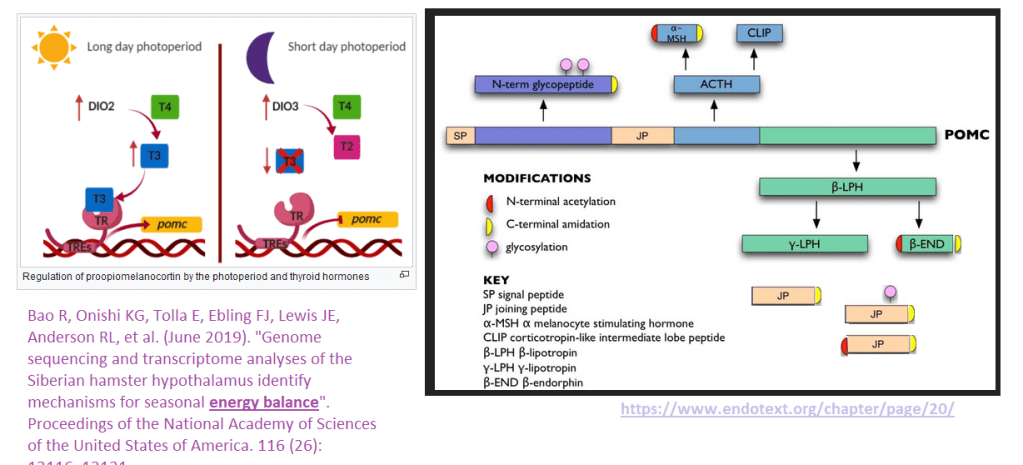
Pro opiomelanocortin
This gene contains many peptide hormones in one transcript. Then a protease comes and cleaves the new protein into many peptide hormones.
Left Iodothyronine deiodinase 2 rohormone thyroxine (T4) to the active hormone triiodothyronine (T3) via an iodine removal such that T3 can bind to the hormone receptor (TR), which then binds to thyroid hormone response elements (TREs) in the promoter of the POMC gene. In periods of less sunlight, there’s a conversion to an inactive form of thyroid hormone. See also: Barrett P, Ebling FJ, Schuhler S, Wilson D, Ross AW, Warner A, et al. (August 2007). “Hypothalamic thyroid hormone catabolism acts as a gatekeeper for the seasonal control of body weight and reproduction”. Endocrinology. 148 (8): 3608–17. doi:10.1210/en.2007-0316. PMID 17478556 Right: The POMC gene. Only a few of the peptide products are C-terminal amidated, yellow cap. The original image was modified based on dta in reference [9]
A short term copper deficient human study…
A 1998 human study investigated the effect of a low copper diet on plasma opioid peptides. In this study in 11 healthy young volunteers were fed foods low in copper but adequate in all other nutrients. [7]
0.66 mg/day for 24 days (marginal Cu)
0.38 mg/day for 42 days (low Cu) ↓ ceruloplasmin & plasma Cu, ACTH
2.49 mg/day for 24 days (adequate Cu). ↑ ceruloplasmin & plasma Cu
TPlasma β-endorphin, Leu-enkephalin, Met-enkephalin, and Adrenocorticotropic hormone (ACTH), as measured by immunoassay, did not significantly change between the start and finish of each study period. [8] intake.
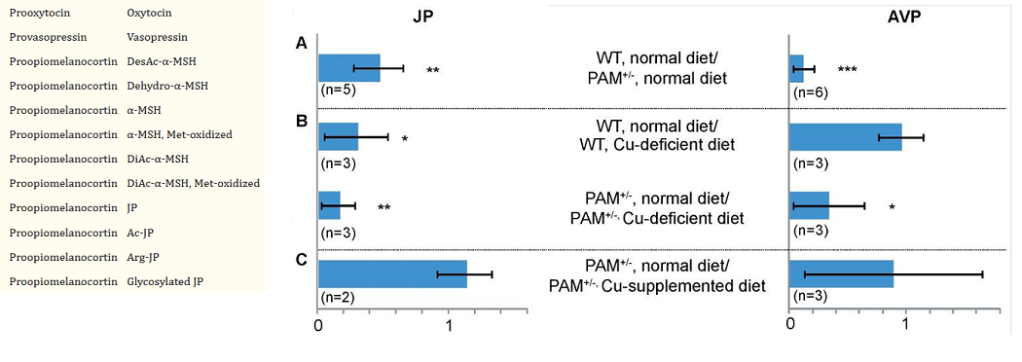
PAM+/- mice and a mass spectrometry technique
Yes ground up pituitary glands were involved in these experiments. Not only were the masses of these peptides measured with mass spectrometry, but also the masses of the fragments of busted up peptides. This allowed the investigators to detect new modifications of the peptide hormones. Recall that JP is the joining peptide between
ManyC-terminal amidated peptides were detected. Just a few showed statistically significant change. It has been hard to pin down a function for the joining peptide. More interest seems to be in AVP in terms of cardiovascular health than emotional health.
Where PAM is expressed
This image was derived from proteinatlas.org. The expression of PAM in the brain is limited. It is assume that this image from Protein Atlas is from the atrium.
In spite of PAM being involved in production of “fee good”hormones, it’s most abundantly expressed in the heart and a few other organs we are interested in This particular immunocytochemistry image of the heart from Protein Atlas looks like PAM is in some sort of vesicle. In spite of its abundance in the heart, its function is kind of hard to pin down. mRNA data indicates that PAM is comparatively abundance in the hypothalamus of the brain. The heart expression is kind of hard to ignore. There is an interest in measuring secreted PAM as an indicator of heart disease. A 96 well plate assay for detecting PAM activity in the serum has been developed by a company called Pam-t.com. [10]
References
Bousquet-Moore, D., Mains, R. E., & Eipper, B. A. (2010). Peptidylgycine α-amidating monooxygenase and copper: a gene-nutrient interaction critical to nervous system function. Journal of neuroscience research, 88(12), 2535–2545. PMC free article
Bousquet-Moore, D., Prohaska, J. R., Nillni, E. A., Czyzyk, T., Wetsel, W. C., Mains, R. E., & Eipper, B. A. (2010). Interactions of peptide amidation and copper: novel biomarkers and mechanisms of neural dysfunction. Neurobiology of disease, 37(1), 130–140. PMC free article
Lee, H. J., Macbeth, A. H., Pagani, J. H., & Young, W. S., 3rd (2009). Oxytocin: the great facilitator of life. Progress in neurobiology, 88(2), 127–151. PMC free article
Prohaska, J. R., Gybina, A. A., Broderius, M., & Brokate, B. (2005). Peptidylglycine-alpha-amidating monooxygenase activity and protein are lower in copper-deficient rats and suckling copper-deficient mice. Archives of biochemistry and biophysics, 434(1), 212–220. PMC free article
Yoshimura, M., Conway-Campbell, B., & Ueta, Y. (2021). Arginine vasopressin: Direct and indirect action on metabolism. Peptides, 142, 170555. PMC free article
Song, Z., & Albers, H. E. (2018). Cross-talk among oxytocin and arginine-vasopressin receptors: Relevance for basic and clinical studies of the brain and periphery. Frontiers in neuroendocrinology, 51, 14–24. PMC free article
Bhathena SJ, Werman MJ, Turnlund JR. Opioid Peptides, Adrenocorticotrophic Hormone and Dietary Copper Intake in Humans. Nutr Neurosci. 1998;1(1):59-67.
Yin, P., Bousquet-Moore, D., Annangudi, S. P., Southey, B. R., Mains, R. E., Eipper, B. A., & Sweedler, J. V. (2011). Probing the production of amidated peptides following genetic and dietary copper manipulations. PloS one, 6(12), e28679. PMC free article
Kaufmann, P., Bergmann, A., & Melander, O. (2021). Novel insights into peptide amidation and amidating activity in the human circulation. Scientific reports, 11(1), 15791. PMC free article
We at CopperOne think that any copper in the cuprous, Cu+, oxidation state is best because entering through the specific Ctr1 channel is like entering a hotel through the front door. Once you enter, you have a bell hop and concierge to help you find your way to your room and make sure that electron you’ve got in your outer valence shell doesn’t go where it shouldn’t. Like O2!!!
Only copper in the Cu+ oxidation state is allowed to enter the hotel of the cell through Ctr1.
DMT1 is like a side door
We at CopperOne call the +2 oxidation state Cu2+ “toxic copper.” Well, it is better than nothing if one is a cow or goat eating copper deficient vegetable matter and made more copper deficient with molybdenum. (For a very technical post, follow this link.) We hear stories of humans taking mega doses of copper sulfate. Why??? Sure, it can enter through the divalent metal cotransporter (DMT1) side door. Why should some of the other divalent minerals that our bodies need have to stand in line?
These are some of the divalent metal cations that
Sure, Cu2+ entering through the DMT1 side door will eventually get reduced to Cu+ via glutathione or something. Not that this is speculation waiting to be experimentally verified. It will make its way to its room, we hope. It just seems profoundly ill advised to take mega doses of any form of Cu2+… to us anyway.
It should be noted that both Cu+ and Cu2+ can bind to the transcription factor Sp1 to shut down the expression of Ctr1.
This post examines a report from the laboratory of Dr Christopher Fahrni at Georgia Tech University. What this group is saying is that the small molecule glutathione can chaperone intra cellular Cu+ to really low femptomole levels. [1]
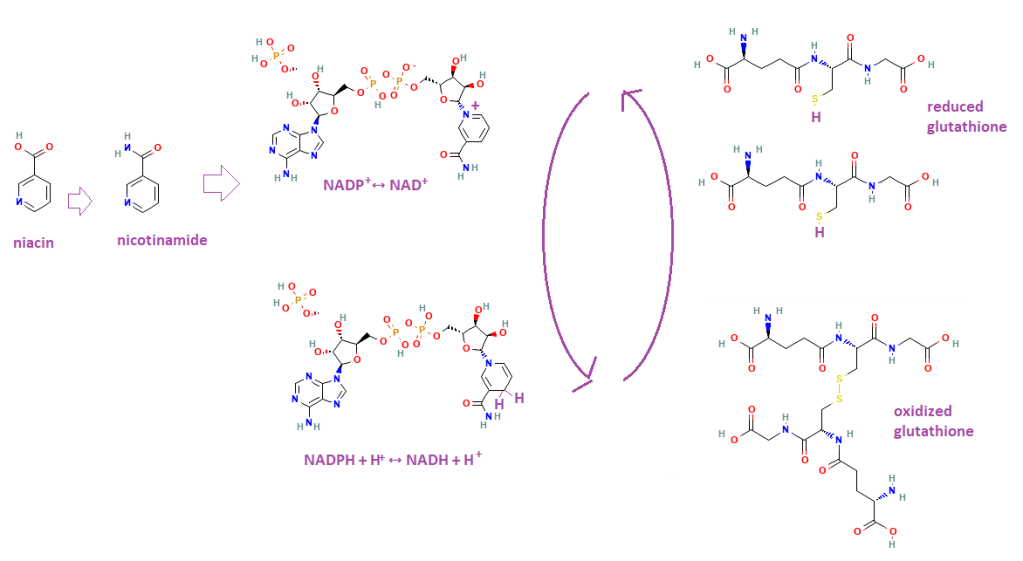
This is a glutathione molecule. The carboxy C-terminus (had end) is a glycine. A carboxy group is -COOH. The “butt end” N-terminus of glycine is –NH2, an amino group. The middle amino acid is cysteine with the yellow –SH side chain. The Carboxy group of csyteine can come behind the butt end of glycine and take a big ole bite and kick off a H2O in the process. An amide bond is formed. The tail end of glutathione is formed when the -COOH of the side chain of glutamine comes in and takes a big ole bite of the butt of cysteine. Again, a H2O is formed from the OH of the glutamine side chain and a Hof the -NH2 amino terminus of cysteine. The The pKa values are from UCLA.
a molecule of glutathione (GSH) . Note, biochemists call the butt end the N-terminus. The “head” end is called the C-terminus.
The pKa, to be protonated or not
The pKa in very simple terms is the pH in which 50% of the green circled “H” are there are dissociated leaving no charge in case of the amino (butt) terminus -NH2 group or the carboxy terminus -COO– group.
The official macroscopic pKa values corresponding to sequential deprotonation of glutathione are as follows:
the Glu-α-COOH, 1.98
Gly-COOH, 3.49
–SH, 8.64
Glu-NH3+ , 9.36.
with the note that GSH exists primarily as a mono anion at neutral pH. [1]
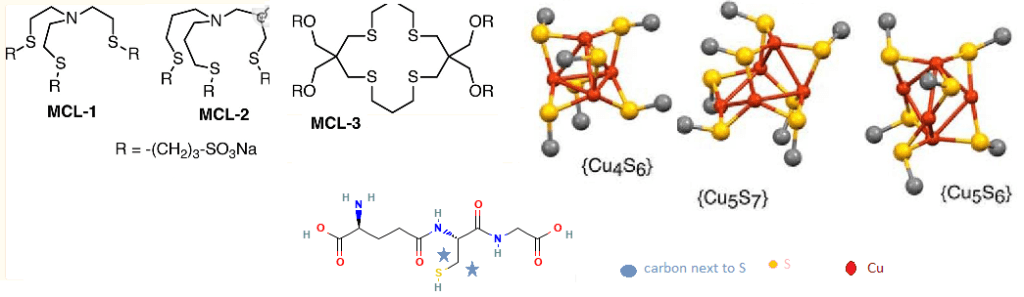
The Fahrni group used three water soluble monovalent Cu ligands (MCL1-3) with known affinities for Cu+ to compete with GSH. We will not present the sophisticated optical techniques they used to use these competing MCL1-3 copper ligands to measure the interaction of GSH molecules for Cu+.
the concept of glutathione clusters
Quoted from the publication:
“Free GSH exists primarily in mono anionic form at pH 7, whereas cluster formation presumably entails S-deprotonation to the corresponding dianion, which we represent as GS−. An equilibrium between clusters differing in Cu(I)/glutathione ratio would therefore depend on [GS−], which is influenced by both [GSH] and pH. Once the latter is significantly below the thiol pKa of 8.75, each further unit drop in pH will decrease the GS−/GSH ratio by 10-fold. We therefore measured pairs of solutions containing either 1 mm GSH or 10 mm GSH at 1 unit lower pH.” The typical GSH-Cu complex was determined to be [Cu4(GS)6] as the dominant species over a wide pH range, from 5.5 to 7.5. [1]
MCL1-3 are probes the group used to compete with GSH for Cu+. Cu4S6, Cu5S7, and Cu5S6 are theoretical models based on other studies in the literature. Only the carbon next to the Cu binding S is shown for clarity.
Skipping some complicated techniques, the Fahrni group came up with an equation for “parts” Cu+ that is analogous to parts H+, or pH. pH is the –log base 10 of the concentration of H+ in molar. The log10 of 0.1 M H+ is “-1” Minus the log10 is pH 1. Going in the opposite direction, the concentration of H+ at pH 10 is 10-10 moles per liter H+, or 0.0000000001 moles per liter!
Say some catastrophic even occurs and results in 10 mM GSH being oxidized to 4 mM GSSG leaving only 2 mM GSH.
The first term goes from 1.5x -2 = -3 to 1.5x-2.7 = -4
For the 2nd term, say we go from 1.5 x pH 7.4 = 11.1 to 1.5 x acidic pH 6 = 8
10uM total Cu = -0.25x (-5)= 2.5 and 1uM = -2.5x (-6)= 1.5
Good conditions: pCu= -3 + 11.1 – 2.5 + 8.26 = 14 10 -14 M free Cu is really low. We could be at 10-15 M or 1 fM free copper if we went even lower in total copper.
Not so good conditions: pCu = -4 + 8 -1.5 + 8.26 = 10.8 = 10-11 M free Cu. pH and reduced glutathione are far more important determinants of free copper, than total copper.
Prior to the Fahrni group’s study, scientists had known for decades that Cu+, in the very reducing interior environment of the cell, formed complexes with reduced glutahione. This is the genius of cuprous nicotinic acid that we call CopperOne.
Why we need niacin and NADH/NADPH
Niacin is a precursor to nicotinamide, which is part of nicotinamide adenine dinucelotide (NAD) that may be phosphorylated shown as a phosphate not totally attached to a ribose group. The NADPH/NADP of source of reducing equivalents maintains the reduced state of the GSH thiol.
Bernt and coauthors have written a nice review on the role of thioredoxin and glutaredoxin in maintaining the reduced state of glutathione and protein thiols. [2] This figure only shows GSH in a GSSG disulfide bond that prevents proper buffering of intracellular copper. Further oxidation states are also possible. if it were not for NAD/NADP and thio and glutaredoxin. [2]
We become so worried about an increase in total copper in our cells and forget that free copper is more deleterious.
If we have a decrease in electron transport chain activity because complex IV lacks Cu, we have an increase in glycolysis and a decrease in pH. This increases free Cu. If we do not have fully active Cu/Zn superoxide dismutase, the same -S group in GSH might also become oxygenated to cysteine sulfenic acid -SOH that is a precursor to -S-S- disulfide bond. [2] Higher order cysteine oxidation products are possible. [2] Perhaps we should worry less on the affect of extra copper on free copper when we also should think about the role of niacin and NAD that contribute to this remarkable balance of nature that keeps the free copper in our cells insanely low!
References
Morgan, M. T., Nguyen, L., Hancock, H. L., & Fahrni, C. J. (2017). Glutathione limits aquacopper(I) to sub-femtomolar concentrations through cooperative assembly of a tetranuclear cluster. The Journal of biological chemistry, 292(52), 21558–21567. PMC free article
Berndt C, Lillig CH, Holmgren A. (2007) Thiol-based mechanisms of the thioredoxin and glutaredoxin systems: implications for diseases in the cardiovascular system. Am J Physiol Heart Circ Physiol. 2007 Mar;292(3):H1227-36. free article
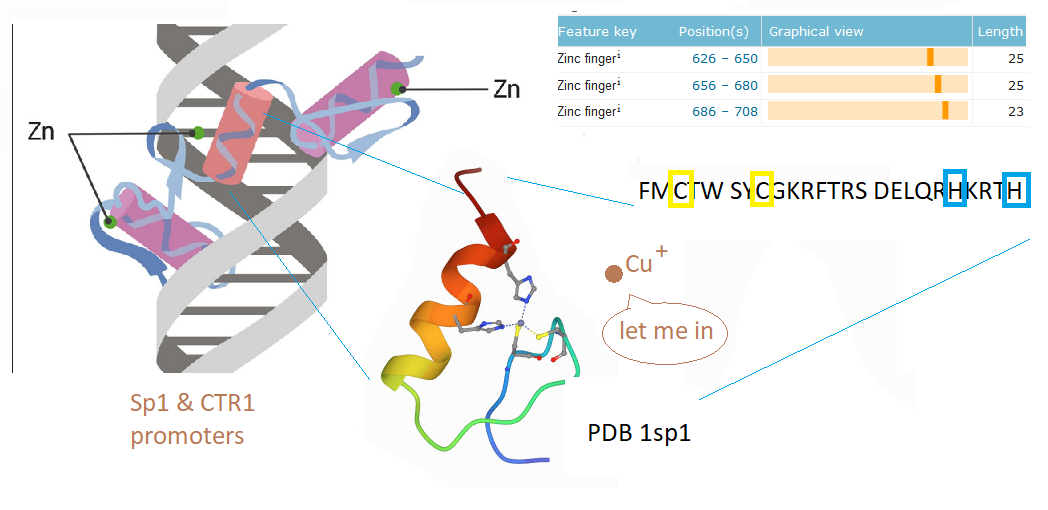
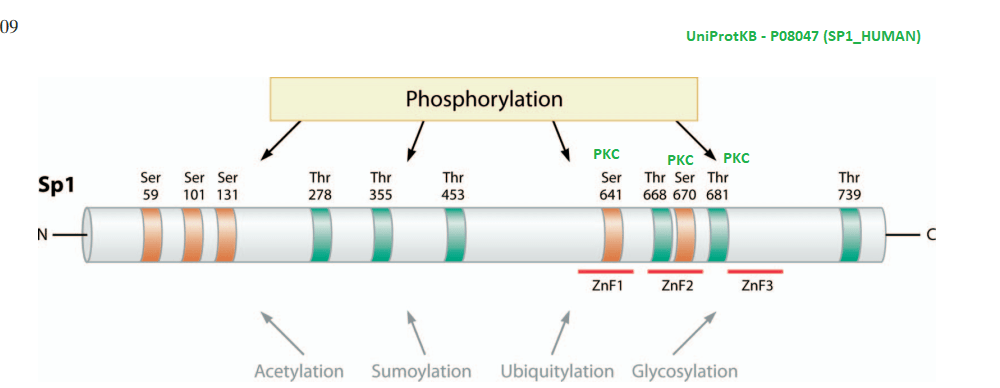
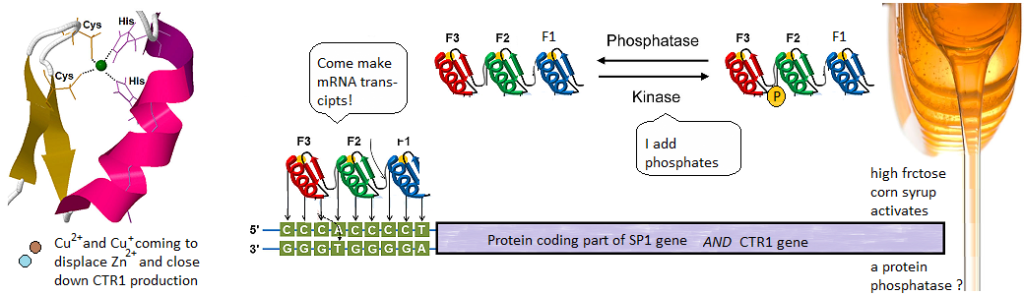
This post investigates a claim in a review by Morrel and coworkers that excessive dietary fructose can decrease the expression of Ctr1 in the duodenum. [1] How could this happen? What is transcription factor for Ctr1? As it turns out, both Cu+ and Cu2+ can bind to the CTR1 transcription factor Sp1 and negatively regulate its activity [2,3] The featured image describes out Cu can disrupt the Zn2+ finger(s?) Sp1 uses to bind to the promoters of itself and CTR1. If you wish to skip the details of some brilliant experiments, click to go to the lay friendly summary.
Cu2+ prevents Sp1 promoter binding
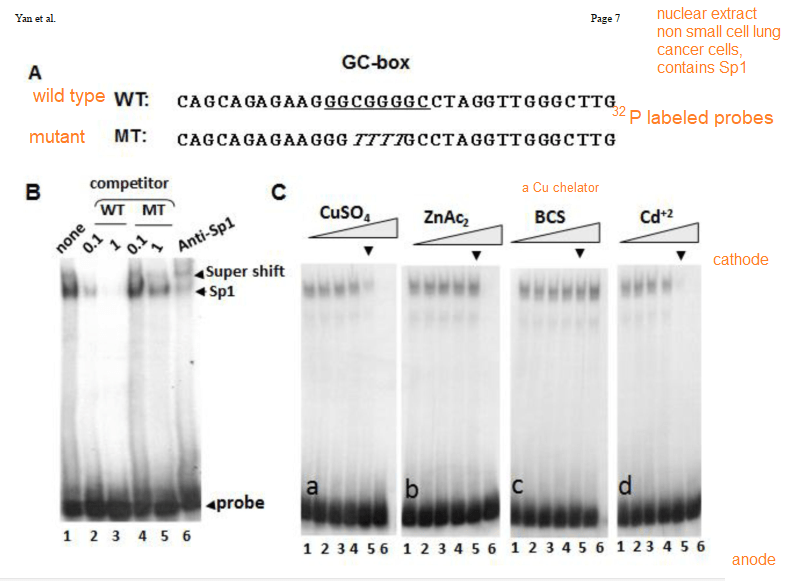
In their introduction to their Cu2+ study Yan and coworkers discussed the role of cysteines and histidines in binding Zn2+ and other transition metals like cadmium and copper. When metal ions are not bound to the zinc fingers of Sp1, Sp1 binds to the GC-rich boxes with the consensus sequence 5’-G/T-GGGCGG-G/A. This GC box lies upstream of the gene for Sp1 itself, CTR1, and many other genes. In the absence of copper, Sp1, the protein transcription factor, increases transcripts for more Sp1 and Ctr1 until copper balance is restored. Figure 1 of this study used a nuclear extract of non small cell lung cancer cells that contained several isoforms of the SP1 protein. This extract was mixed with 32P labeled double stranded DNA probes that have a negative charge. The mixture was added to an agarose gel. An electrical current was applied such that the negatively charged small pieces of probe DNA migrated to the anode. When bound to Sp1, they migrated slower. Concentration ramps of t 0, 1, 10, 50, 100, and 200 µM divalent metal ions were added to test the hypothesis that less probe would be present in the upper band that contains Sp1. A Western blot was performed on the material in 1B to demonstrate that the upper band contains Sp1.
The first lane of 1B is just the 32P labeled probe. The second two lanes are two increasing concentrations of the wildtype probe that is not32P labeled. The two MT mutant probes are also not radioactive. These results demonstrate the specificity of the assay. [2]
Cu+ prevents Sp1 promoter binding
Technically speaking, there should be no free copper inside the cell. It should be bound to some chaperone. Cu should be in the Cu+ oxidation state. This is what is ground breaking in the Yuan 2017 study. Sp1, even when its Zn2+ fingers are metallated and ready to bind to CTR1 promoter, the chaperone Atox1 can donate a Cu+ and turn it off.
1 Cu+ binds to all three Zn fingers
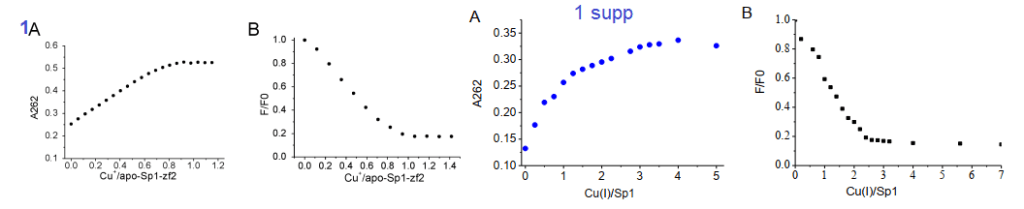
Yuan and coworkers kept Cu in the +1 oxidation state by use of the reducing agent TCEP. The absorbance at 262 nm was taken as an indication of S- Cu+ charge haring transition. [3] This particular experiment only used the second Zn2+ finger. The second Zn2+ finger contains two tryptophans W560 and W571. Tryptophan fluoresce at 340 nm when excited at 280 nm. Fluorescence may be quenched when these aromatic residues are exposed to an aqueous environment, or as the authors speculated, by interactions of the thiolate anion with Cu+. The authors also stated that the apo 2nd Zn2+ finger tends to be unfolded. The amino acid sequence of the 2nd Zn finger is shown in the featured image note that there’s a tryptophan (W) flanked by two cysteines ( C yellow box).
1A the second zinc finger from Sp1. !supp, the full length Sp1
Note that Cu binding data for just the 2nd Zn finger saturate at 1:1 Cu per Sp1. For the full length Sp1, the binding saturates at a 3:1 ratio. This suggests that all three Zn fingers bind Cu.
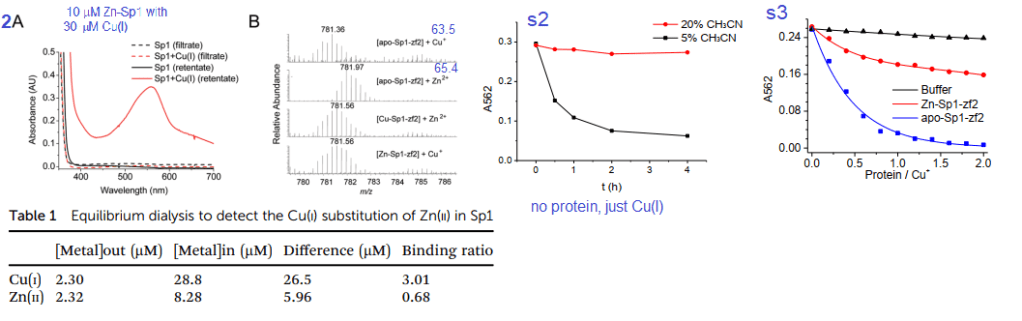
2. Cu(I) can react with Zn-bound Sp1
Figures 2A, s2 and s3 used a Cu+ chelating agent called bicinchoninic acid (BCA). BCA has a strong absorbance maximum at 582nm when it binds to Cu+. Panel 2A is not (just) the absorbance spectrum of Sp1 with Cu+. BCA was added to the material retained on an ultrafiltration membrane with a 3 kDa cutoff size. Anything smaller is passed through the filter and larger is retained in a small amount of solution on the top of the filter. As a negative control, the filtrate and retentate of Sp1 that had never seen Cu did not react with BCA to give the magenta color. Table 1 shows the results. Almost all of the 30 μM Cu+ is picked up by 10 μM Sp1. About 2.μ. There was 2.32 μM Zn2+ displaced from 10 M Sp1. 8.28 Zn2+ remained “in†the dialysis tube with the Sp1. These data estimate the Zn2+ – Sp1 binding ratio to be 0.68 and 3.01 for Cu+ to Sp1.
Panel 2B used a technique called mass spectrometry to measure the mass/charge ratio of the Sp1 2nd Zn finger. There is a semi clear reduction in mass when Cu+ displaces Zn2+ These two atoms are very close in mass. To make matters more interesting histidines and cysteines have side chains that may be protonated, or not depending on the pH of the solution and whether they are binding transition metals. Supplemental figure 2 shows use of 5 and 30% acetonitrile as a means of slowing down oxidation of Cu-BCA. Supplemental figure 3 shows just the 2nd Zn finger of Sp1 stealing Cu from CuBCA in the presence of 10% v/v acetonitrile. In just buffer, the absorbance stays fairly constant. As Zn-Sp1-zf2 is added, The absorbance of Cu-BCA decreases about one third of the original. When there is no Zn, the decrease in Cu-BCA absorbance is essentially complete when there is twice as much 2nd Zn finger as BCA bound Cu. Just a note, UV /visible light absorption is often used to measure metal ions interacting with proteins. These contributions are probably not that great in this particular system.
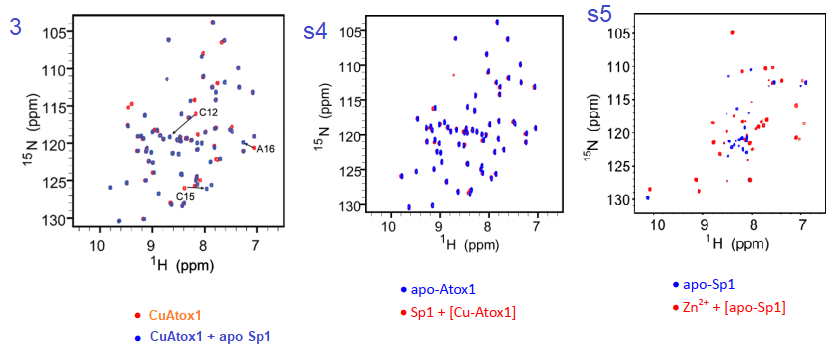
3. NMR experiments
The authors did not disclose details of how they labeled the expressed protein with 15N instead of the more abundant 14N isotope. It is assumed that they supplemented the bacterial expression system with 15N ammonium chloride or something. The NMR technique has heteronuclear single quantum coherence (HSQC) that basically measures 1H-15N pairs within the amino acids of the protein. Each dot on the graph represents a bonded H and N in only the 15N labeled protein.
Figure 3 and complementary supplemental dtaa from Yuan (2017) [3]
Yuan and coworkers reported that the association constant of Cu for KCu-Atox1 = 2.51x 1017protein. Adding Apo Sp1 changes a number of N-H pairs. Figure s4, going in the reverse direction changes little. We are assuming that Cu-Atox1 is the 15N-labeled protein. Figure s5 is somewhat of a control showing some massive, global changes when Zn2+ binds apo Sp1. Note that these changes seem to be larger than when Cu+ binds apo Sp1.
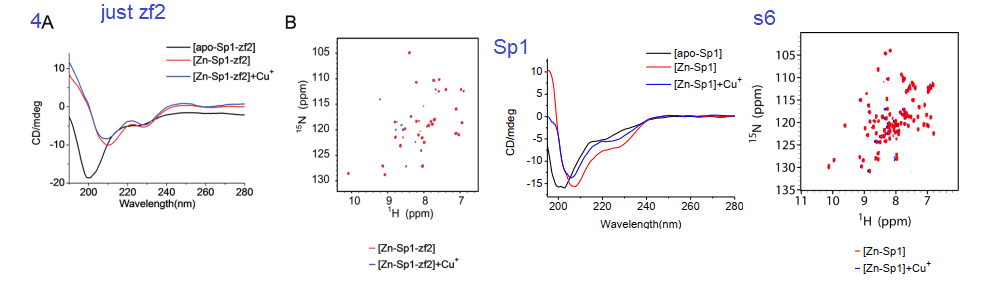
4. Circular Dichroism, a window into protein secondary structure
Circular dichroism is a method involving the absorbance of circularly polarized light by structures within a protein. It is used to measure the proportions of α-helices, β-sheets, and random structures. The yellow to red helix in the featured image is considered an α-helix. Binding of a metal ion may be anticipated to make a protein more ordered.
Fig 4 and supplemental from Yuan 2017
Yuan and coauthors considered that the addition of Cu+ and Zn2+ both resulted in a more ordered zf2. The 2D HSQC NMR data in 4A and s6 also indicated not that much change between the two metal ions and the existence of a Cu finger. Figure s6 is a mirror experiment using the entire Sp1 protein. Zn-Sp1 + Cu+ seems to be somewhere in between the disordered apo Sp1 and Zn-Sp1 structures. Cu+ disruption of GC box binding is what really matters in this story.
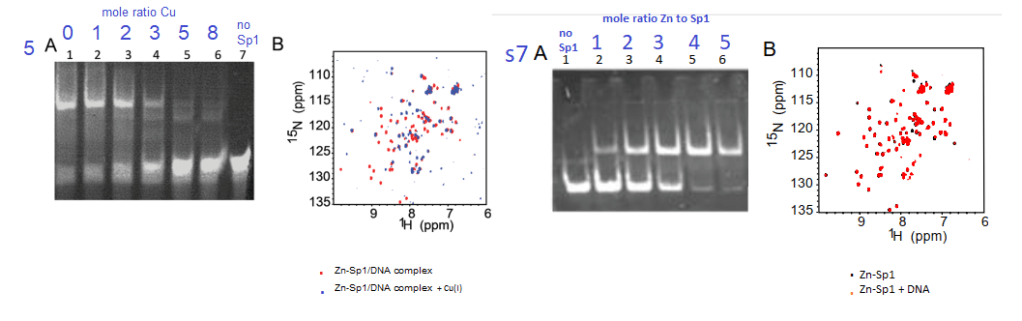
5 Cu+ inhibits Sp1 from binding to GC boxes
Yuan and coworkers also used the gel mobility shift assay to demonstrate tha at about 3:1 moles Cu+ to 1 mole Sp1, the probe of the GC rich binding site on the CTR1 promoter ceases to bind Sp1 (top band, panel 5A. Panel s7A shows the opposite is true for Zn2+. These authors used a lot of NMR data
Figure5 and supplemental figure 7 from Yuan (2017)
What about post translational changes in Sp1 other than metal binding? Tan and Khachigian wrote an insightful review in this regard.[4]
A highly edited Table 1 from Tan and Khachigan 2009. The many phosphorylation events that lead to activation of Sp1 have been removed.
Some further investigation of the cited references revealed hexose kinase was also down regulated by glucose deprivation. [5] The PP1 inhibitor okadaic acid decreased the increased transcription of aldolase and pyruvate kinase in response to glucose. [5] These authors saw what they interpreted as minor proteolysis of the dephosphorylated Sp1 in their Western blots. [5]
Note that two protein kinase C phosphorylation sites flank Zn finger #2. Protein kinase C is activated by Ca2+.
Concluding remarks
We are little closer to understanding how fructose can lead to copper deficiency than we first began. Over dosing on Cu is still possible. It would seem that nature has given us a way to turn off import when we have enough. Sp1 even “knows” when our Cu chaperones are copper replete. [3] We are only scratching the surface of possible Sp1 post transnational modifications that might impact Sp1 binding to the CTR1 promoter. It is becoming apparent that the cure for copper deficiency may not be as simple as eating more copper.
S-glutathionylation is a postranslational modification that occurs during oxidative stress whereby glutathion forms S-S bonds with protein thiols. Recall that the SH group of Zin finger cysteines. Glutathionylation may be reversed with NADH dependent enzymes thioredoxin and glutaredoxin.
Advanced glycation end products. AGE are formed when -NH2 on side chains of amino acids such as lysine react with the aldehyde groups of sugars, fructose more than glucose. Note that there are a few lysines (K) in the sequence of the 2nd Zn Finger of Sp1.
N-acetylation of lysines may occur on K2 and K703 of human Sp1 according to UniProt.org. K703 is within the 3rd Zn finger. We at CopperOne think that a copper replete, functioning mitochondria is needed for generation of NAD+ which regulates the deacetylation enzyme Sirt1.
Lay_Summary
Sp1 binds upstream of the protein coding parts of its own gene and the gene for the Ctr1 transporter. It contains three Zinc fingers. Cu+ and Cu2+ can displace the Zn and stop Sp1 from making more Sp1 and Ctr1. Sp1 can have phosphates attached to it by enzymes called kinases. Phosphatases take phosphates off. Phosphates some how or another may increase transcription of down stream genes. Speculation is that fructose may activate a phosphatase.. We’ve still no clue as to how fructose causes copper deficiency… just some educated hypotheses.
For CopperOne critics who don’t like us using Dr Brewer as a reference, we are remaining fast in our conviction that Cu2+ is bad. A new colleague shares this conviction. If Cu2+ is absorbed via the divalent metal ion transporter, the right way to absorb proper Cu+ gets shut down.
References
Morrell, A., Tallino, S., Yu, L., & Burkhead, J. L. (2017). The role of insufficient copper in lipid synthesis and fatty-liver disease. IUBMB life, 69(4), 263–270. PMC free article
Yuan S, Chen S, Xi Z, Liu Y. Copper-finger protein of Sp1: the molecular basis of copper sensing. Metallomics. 2017 Aug 16;9(8):1169-1175. PMC free article
Yan, D., Aiba, I., Chen, H. H., & Kuo, M. T. (2016). Effects of Cu(II) and cisplatin on the stability of Specific protein 1 (Sp1)-DNA binding: Insights into the regulation of copper homeostasis and platinum drug transport. Journal of inorganic biochemistry, 161, 37–39. PMC free article
Tan, N. Y., & Khachigian, L. M. (2009). Sp1 phosphorylation and its regulation of gene transcription. Molecular and cellular biology, 29(10), 2483–2488. PMC free article
Schäfer D, Hamm-Künzelmann B, Brand K. (1997) Glucose regulates the promoter activity of aldolase A and pyruvate kinase M2 via dephosphorylation of Sp1. FEBS Lett. 1997 Nov 17;417(3):325-8. free article
This review carried the torch of Dr Brewer in trying to shed light on the hazards of using copper (II) sulfate to control pathogens on food crops to the international community. This review cited the literature on links to Alzheimer’s Disease and alternatives to CuSO4 to control pathogens.  This review did not discuss whether the CuSO4 used to control pathogens gets incorporated into nicotianamide or some other less toxic compound.Â
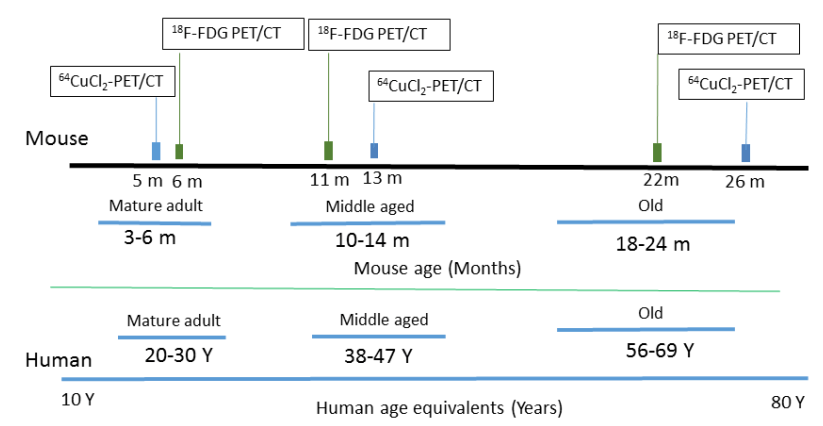
The use of 64Cu PET imaging is continuing to teach us things about how copper moves about mammalian bodies, in particular the brain. This particular study was a collaboration between UT Southwestern and Wayne State. Production of the tracer 64CuCl2 required a biomedical cyclotron and 64Ni(p,n)64Cu at Washington University (St Louis, MO) A 0.1M HCl solution was used to dissolve the radioactive tracer. The goal was to follow 64CuCl2 orally ingested in a common strain of mice, C57BL/6 mice, at various ages.
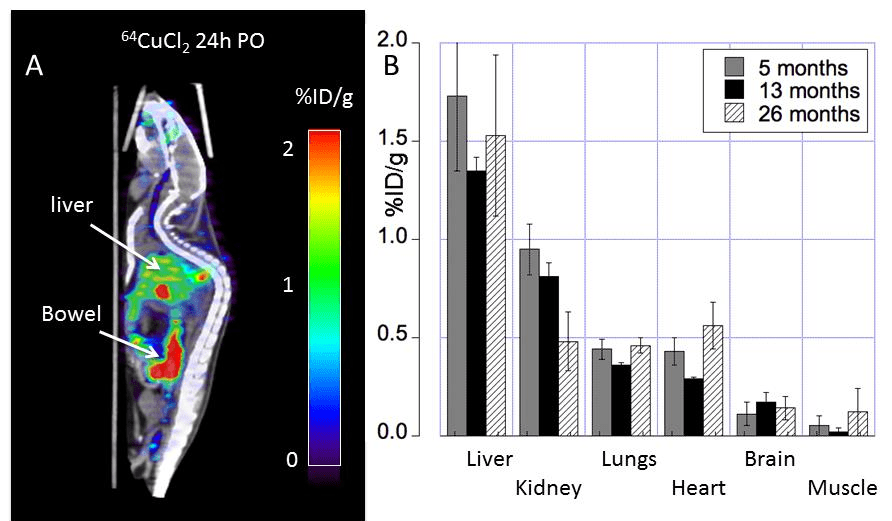
Figure 1 from Peng 2018. Note that both the copper and glucose analog FDG were given orally.
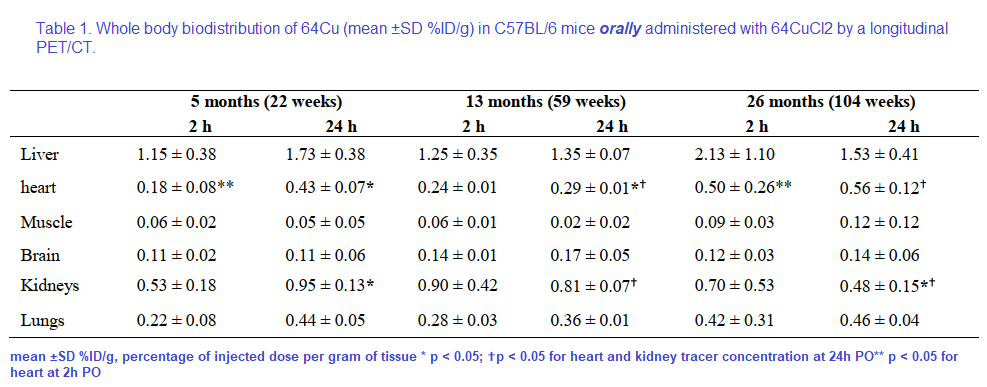
18F-FDG, or 2-deoxy-2-[F-18]-fluoro-D-glucose is used to visualize where the glucose is going in the circulation. What makes this study unique is the oral dosing, 64CuCl2 (2 μCi(74 kBq)/g body weight) diluted in normal saline. Delivery was via a blunted oral feeding tube, followed 15min static whole-body imaging at 24 hours. It was the heart and kidney that showed the greatest age-related differences.
Table 1 from Peng 2018 [2]
Note that the percentages of the initial dose don’t come even close to adding up to 100% Figure 2 gives us a reason as to why.
Figure 1 from Peng 2018 [2]
Note the large amount of the 64Cu is in the bowel. The % initial doses do not seem to add up to be 100%. How much of the initial 64CuCl2 was excreted in the urine or feces? Are the blue speckles underneath the red labeled object labeled “Bowel†feces? The liver is green with yellow horizontal stripes. Is this indicative of 64Cu being incorporated into a chaperone protein like ceruloplasmin?
The glucose utilization does not seem to follow copper deposition. For some reason muscle and renal utilization seem to increase the most in going from middle to old age.
Table 3 Peng 2018 [2]
In going from middle age to old mice, muscle and kidneys showed the greatest proportional increase. The most telling nuance is the whole animal imaging.
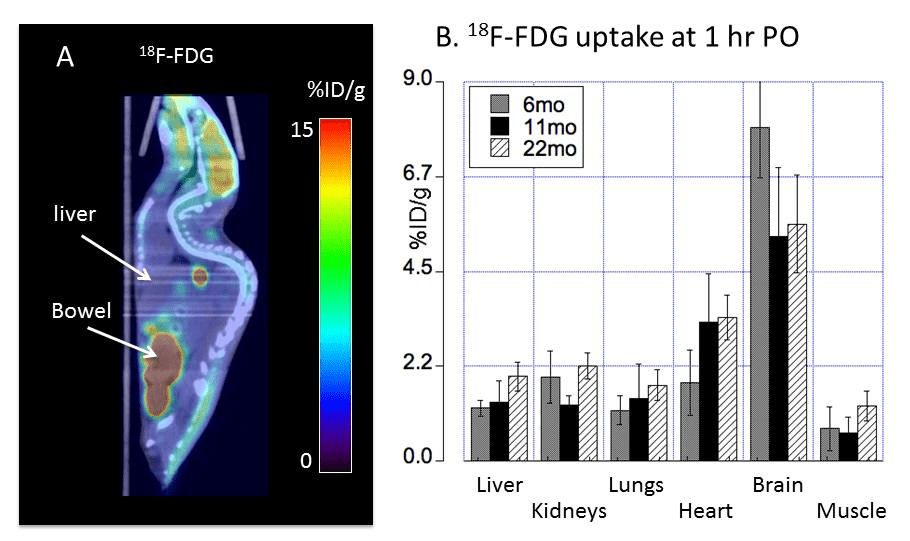
Fig 4 Peng 2018 [2]
The metabolic demands of the liver are fairly high. The liver uptake of glucose might be high when conditions are conducive for production of glycogen. Note that mice were given FDG orally prior to a fast. “Bowel†here likely refers to the small intestine. Also note that there are not little balls going down that would be suggestive of this glucose analog being lost in the feces.
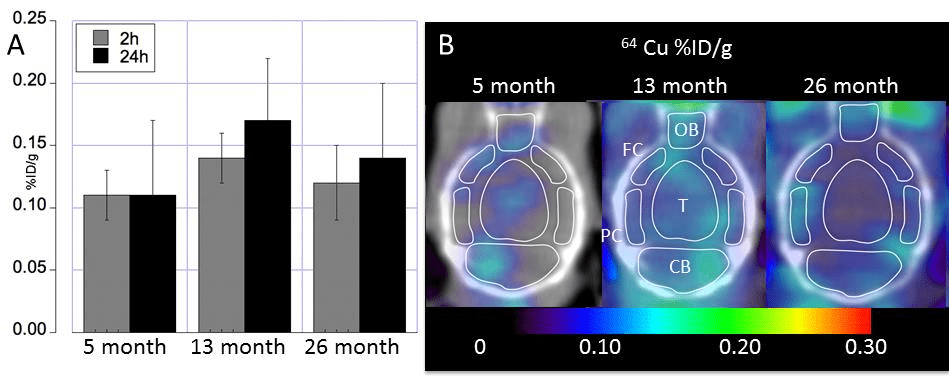
Really no change in Cu uptake in the aging mouse brain?
The apparent no change in the aging mouse brain masks in increase in brain stem and decreases in the hypothalamus/thalamus and basal ganglia.
The FDG measurement of glucose utilization was lower in all regions in the brain in going from 8 to 11 months except the basal ganglia and brain stem. These regions experienced an increase in copper. Is this maintenance of FDG due to maintenance of circulation and dilation of cerebral blood vessels?
Fig 3 from Peng 2018 [2]
Just to round things up, this these are the data for FDG uptake
Table 4 Peng 2018 [2]
It would be interesting to see these experiments repeated with a Copper One supplement like Dr George Brewer recommended.
Cu2+ handling by presenilin
Ca2+ is a natural part of the synaptic cleft function. [3] The presenilin story comes from Ashley Bush’s lab that has long studied how Xu2+ modulates ion channels and what not in the synaptic cleft. This is all a good thing with mechanisms of cleaning up the Cu2+.
Presnilins, clean up proteins and a protease
The γ-secretase is a multiprotein complex that digests integral memembrane proteins. Presenilins 1 and 2 are aspartate protease members of this complex. The amyloid precursor protein (APP) is the most famous target of the γ-secretase complex. When it comes to PSEN1 and PSEN2 mutations that are associated with Alzheimer’s the mutants exhibit altered processing of APP.
Ctr1 is the main importer of Cu+ to the cell, However, 30% of cellular copper import occurs by uncharacteristic mechanisms according to the Bush group’s review of the literature in 2011..The Bush group mentions transporters for Zn2+ and a few of the many enzymes that use Zn2+ as a cofactor. We know that divalent cations like Zn2+ and Cu2+ may catalyze amyloid formation.
Uptake of 0.4MBq of 64CuCl2 (Australian Radiopharmaceuticals and Industrials) and unlabeled CuCl2. Copper was determined in Hank’s balanced salt solution. Uptake was stopped by swashing the cell monolayers of mouse embryonic fibroblasts with ice-cold nonlabeled HBSS (with 2 mM L-histidine).
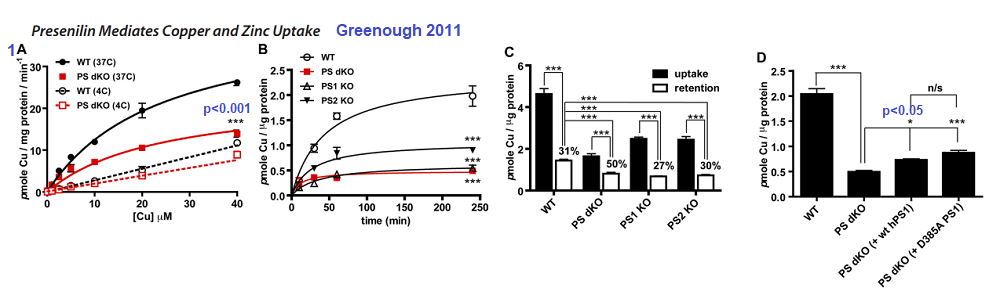
Figure 1 Greenough 2011 [4]
1A The double PSEN1/2 knockout fibroblasts exhibited lower Cu2+ uptake compared to the wildtype at 37oC. The difference was lost at 4oC
1B Time dependent uptake of 2.5 µM 64CuCl2 at 37oC. Loss of PS1 or PS1/2 results in loss of copper uptake.
1C Actual incorporation of copper into proteins is another matter.
1D Rescuing the double Knock Out with human versions of PS1 does not seem to help.
Figure 2 Greenough 2011 [4]
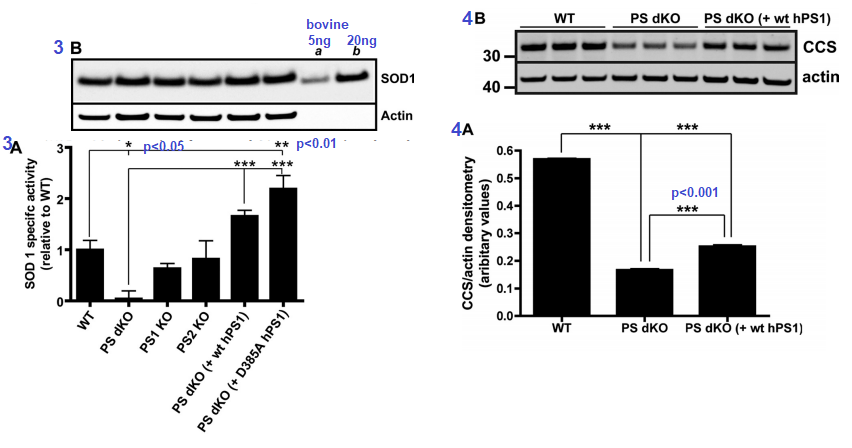
Figure 3 [4] ..Presenilins influence the activity of Cu/Zn SOD1. The assay used mouse embryonic fibroblasts. The D385A mutant of human PS1 is catalytically inactive when it comes to protease activity, but is capable of doubling the Cu/An SOD1 activity. The double knock PS cells had virtually no SOD1 activity suggestive of inability to metalate the enzyme. Expression of SOD1 was fine (3B). Actin is a housekeeping protein used as a loading control.  Â
Figures 3 and 4 from Greenough 2011 [4]
Figure 4 CCS is the copper chaperone for SOD1. CCS is responsible for ensuring that Cu+ is properly inserted into SOD1. Not having both copies of PS translate into not having as much CCS transcribed as mRNA and/or translated into the CCS protein that can be detected in a Western blot (Figure 3B)
Figure 5 looked at Cu2+ and Zn2+ content in soluble and insoluble proteins by tissue, eg brain, kidney, heart… in wild type mice and mice having only one of two copies of the PSEN1 gene. While there as a statistically significant drop from about 65 to 55µg Cu per g soluble protein in the brain, it is uncertain if this difference is functionally significant. Likewise, small but significant decreases in were seen in SOD1 activity and CCS protein amounts in PSEN-/+ mice compared to PSEN+/+ mice, Fig 6.
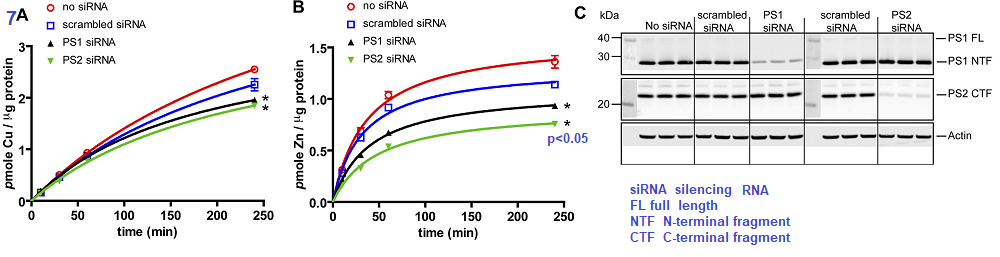
The authors took a likely different approach. They started out with a human embryonic kidney cell line, HEK293T and prevented the mRNA transcripts for PS1 and PS2 from being translated into protein by use of silencing RNA (siRNA) Scrambled siRNA is the same complementary sequences scrambled such that it cannot bind to the untranslated mRNA.
Figure 7 Greenough 2011
The authors demonstrated that silencing of mRNA translation into proteins resulted in a small but significant decrease in Cu loading of proteins (7A) and perhaps a 50% decrease in Zn protein incorporation (7B). Panel 7C shows that the siRNA was successful in knocking down mRNA translation into presenilin proteins. The authors finished up their story in Figure 8 that presented data showing that pharmacological inhibition of protease activity did not affect the Zn and Cu loading.
Concluding statements of the authors brought the reader back to the laboratory’s primary interest: extra-neuronal Cu2+ and Zn2+ released into the synaptic cleft. They cited evidence supporting hyper metallation of A by divalent cations in the synaptic cleft. In glutamatergic synapses, Zn2+ and Cu2+ are uniquely high. [3]
Cu distribution tissue and subcellular
Peng and coauthors demonstrated that Cu2+ distributes distributes differently in young, middle, and old mice. [2] They did not make a big deal of it, but it seemed that most of the copper was being lost in the feces. A parallel study needs to be performed using a more bio available Cu+ source, in our opinion.
Greenough and coauthors made a thoughtful case for presenilin being an alternative way to load proteins with Cu2+ in fibrobasts and in synaptic cleft. [4] These results suggest that nature has a clever way of recycling Cu2+ from the synaptic cleft before it aggregates with Aβ or otherwise cause problems. Why overload this system by drinking water from copper pipes, eating produce treated with CuSO4, or taking cupric dietary supplements.
References
Coelho, F. C., Squitti, R., Ventriglia, M., Cerchiaro, G., Daher, J. P., Rocha, J. G., Rongioletti, M., & Moonen, A. C. (2020). Agricultural Use of Copper and Its Link to Alzheimer’s Disease. Biomolecules, 10(6), 897. PMC free article
Peng, F., Xie, F., & Muzik, O. (2018). Alteration of Copper Fluxes in Brain Aging: A Longitudinal Study in Rodent Using 64CuCl2-PET/CT. Aging and disease, 9(1), 109–118. https://doi.org/10.14336/AD.2017.1025 PMC free article
Opazo, C. M., Greenough, M. A., & Bush, A. I. (2014). Copper: from neurotransmission to neuroproteostasis. Frontiers in aging neuroscience, 6, 143. PMC free article
Greenough, M. A., Volitakis, I., Li, Q. X., Laughton, K., Evin, G., Ho, M., Dalziel, A. H., Camakaris, J., & Bush, A. I. (2011). Presenilins promote the cellular uptake of copper and zinc and maintain copper chaperone of SOD1-dependent copper/zinc superoxide dismutase activity. The Journal of biological chemistry, 286(11), 9776–9786. PMC free article
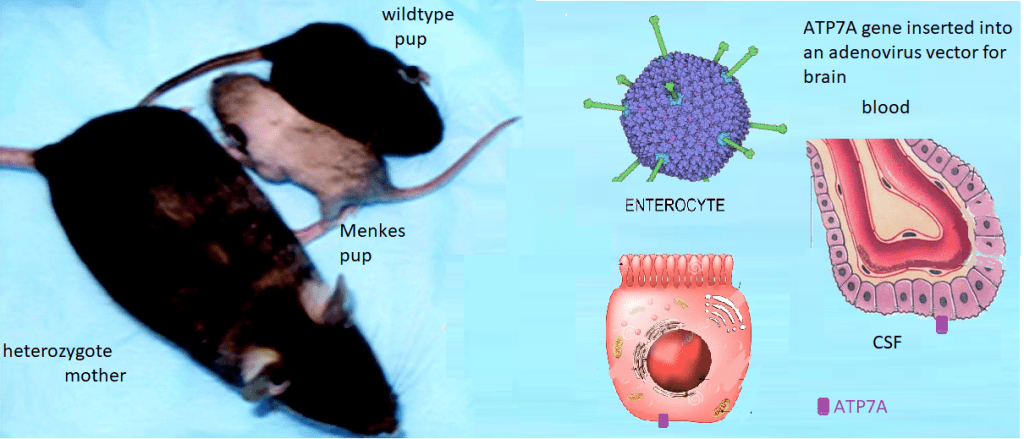
Menkes Disease is an X-linked infant onset disorder of copper transport caused by mutations in the ATP7A copper transporter. [1] Seizures, hypotonia, gross motor delays were some of the neurological deficits caused by mutations in ATP7A. The authors of Wikipedia list a series of symptoms of Menkes Disease that reads like a list of every cuprous enzyme in the body. Steven Kaler’s group describes a somatic mutation in ATP7A that occurred during embryonic development such that not every organ system was affected. [1]
Lay summary of Stephen Kaler’s work. Mice without functional ATP7A copper transporters cannot transport Cu+ from inside the gastrointestinal enterocyte to the blood. Cu+ goes in the opposite direction in the choroid plexus. ATP7A is needed to get from inside the enterocyte to the CSF where Cu can be delivered to the brain. Dr Kaler has been working on gene therapy targeted to these long-lived cells.
This is the lay summary. Some details of experiments are presented. Some CopperOne thoughts on how cuprous niacin might bypass the need to directly inject copper into the blood because it cannot be transported from inside the enterocytes to the blood.
ATP7A gene therapy plus Cu(II)Cl2 injections
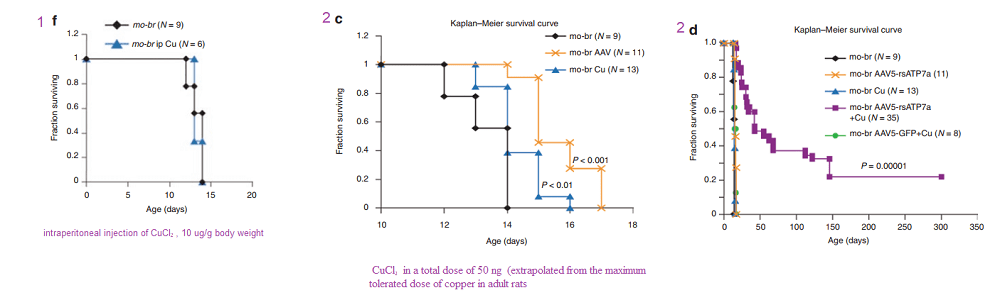
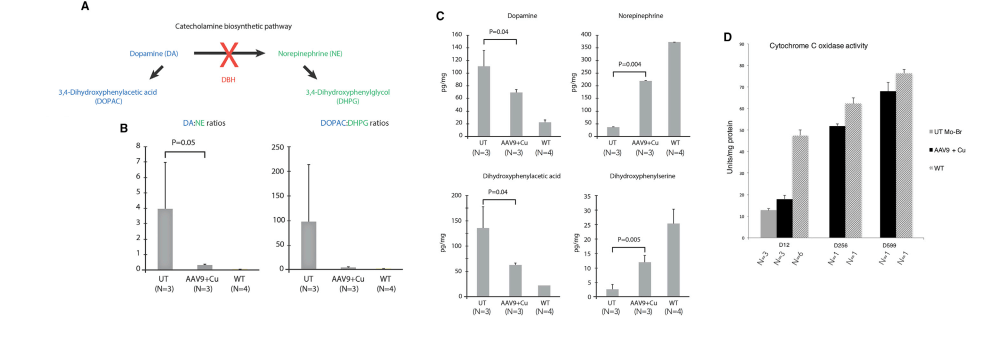
In another Kaler laboratory used a “mottled brown†mo-br mouse mutation at resembles human Menkes disease. Unlike the human treatment that includes subcutaneous injection of copper, the mo-br mutation on C57BL/6 background is not rescued by peripheral copper administration. Neonatal mo-br mice received lateral ventricle injections of either adeno-associated virus serotype 5 (AAV5) harboring a reduced-size human ATP7A (rsATP7A), copper chloride, or both. Brain activity of dopamine-β-hydroxylase, a copper-dependent enzyme, and correction of brain pathology.
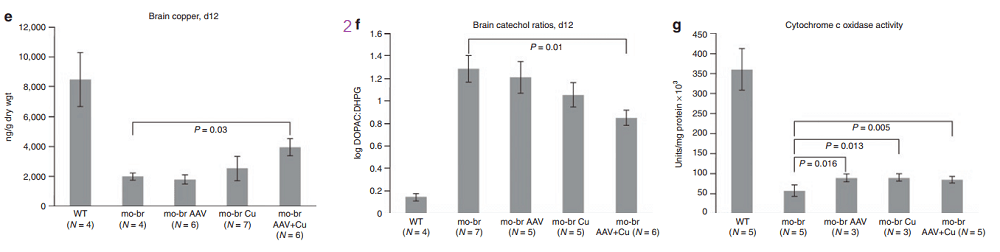
According to the authors, choroid cells are not replaced in the adult lifespan of the mouse. It goes without saying that mice were not were not subjected to intraventicular injection of copper for 300 days. The adenovirus gene therapy was performed on day 2 and the CuCl2 on day 3. Note that only copper treatments extend the brain copper. ATP7A gene therapy was needed to improve copper utilization. Injection of copper and adenovirus ATP7A gene therapy augmented brain copper content. Brain ratios of dihydroxyphenylacetic acid: dihydroxyphenylglycol (DOPAC: DHPG) were increased in untreated mo-br mutants, reflecting dopamine-β-hydroxylase deficiency.
A few years later the Kaler group conducted a similar study with a Cu(II) histidine chelate that is injected subcutaneously. The idea is to bypass ATP7A in the basolateral membranes of enterocytes. These injections improve the outcome with those who ATP7A mutation(s) result n a partially functional proteins.
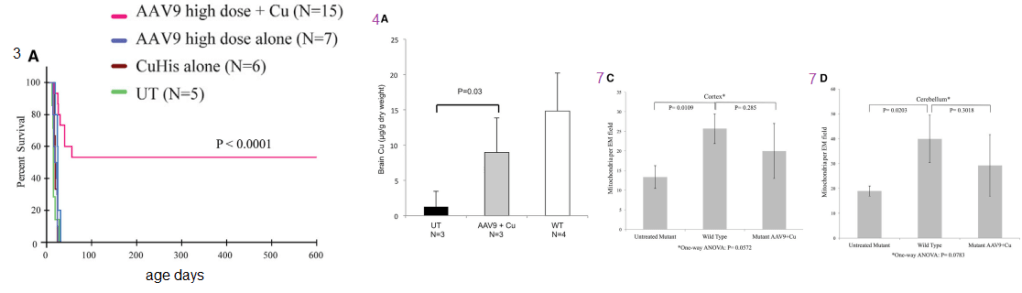
Figure 1 (not shown n this post), the authors performed an adenovirus mock infection with green fluorescence protein and then performed immunohistochemistry to show its distribution. Since this post is about copper, these data will not be presented. Figure 2 showed data validating their transfection methodology and that the reduced size ATP7A responded to increasing copper concentration by moving to the cell membrane. Figure 3 showed some more controls. About 2x as much reduced size ATP7A was expressed in the brains of transfected wild type mice as was full length ATP7A. The survival curve of panel 3A is perhaps most interesting to a copper supplement company.
Panels from Haddad (2018) Figures 3, 4, and 7.
It is not enough to perform ATP7A gene therapy on these mo-br mice. The wild type survive just fine on copper in the rat chow. The gene therapy mice required copper histidinate (1 mg/mL) in a total dose of 15 mg by subcutaneous injection on the back, flank, or neck region, in three 5-mg doses on day 4, day 5, and day 6. This is because they still lack fully functional ATP7A transporters in the basolateral membranes of their GI eptithelial cells. This study went on for 300 days. Figure 3 also showed that the gene transfer was mostly in the brain, with a very low cell turnover. Vectors per mouse genome were less than one in the heart, liver, and muscle. Data were not presented for the small intestine, with a very high cell turnover. Seven of the eight of the Cu histidinate plus AT{7A gene therapy did not make it to day 50 whereas the remaining 8 made it to day 300, Would less have died by day 50 if they had been injected with cuprous histidinate or cuprous glycinate?
What we find interesting about Figure 5 is its relevance to any copper deficiency syndrome that might have causes other than a dysfunctional to marginally functional ATP7A.
Figure 5 from Hadadd 2018 [3]
Tying this back to cuprous nicotinic acid
We have thought of transdermal copper administration. Would an injected or transdermal copper in the +1 oxidation state be more available to Ctr1, see next image, and therefore more able to restore normal neurotransmitter levels?
Some transporters we think about
Monocarboxylate transporters MCT are primarily known as transporters of lactate, a byproduct of glycolysis. Lactate is transported by the blood back to the liver where it is converted to pyruvate and then back to glucose. MCT1 was observed in the apical cytoplasmic membrane of some epithelial cells in the choroid plexus MCT4 was found in the basolateral cytoplasmic membrane of small number of epithelial cells. [4] A recent PET study with the MCT1 MCT1 inhibitor AZD3965 demonstrated that MCT1 is transporter for 11C labeled niacin more so in the kidney, heart, and liver than the brain [5] A 1979 traced the CSF, choroid plexus, and brain appearance of 14C nicatinamide and niacin injected intravenously in rabbits rabbits. [6] Nicatinamide, but not niacin, rapidly entered these compartments. A 2007 review echoed these results that the nicotinamide vitamer ofr niacin is what is transported across the blood brain barrier. [7] That these unnamed transporters are low specificity, high capacity, and unidirection with the provision that nicotinamide tends to be rapidly converted to NAD. [7]
A mass spec proteomics study
Choroid plexuses were isolated separately from the right-lateral, left-lateral, third, and fourth ventricles of 6 month old porcine brains. [8] Note that this study also used leptomeninges in addition to the choroid plexus: left, right, 3rd, and 4th ventricles. [8] A method was used to separate CSF (aptical) and blood (basolateral (blood) membrane vesicles of the meninges. MDR1 and OAT1 were considered as blood (dura)- and CSF-facing plasma membrane markers at the BAB. [8] This figure has been modified from Uchida 2020 [8}: It is presumed that apical and bsaolatreal orientations of the transporters are the same in the choroid plexus as they are in the meninges. Ctr1 and ATP7A (starred) have been added. Relative expression of these transporters has been totally neglected in this post.
Adapted from Uchida 2020 Figure 2.
MCT1, the monocarboxylate tansporter, is involved in transporting, lactate, pyruvate, and branched chain amino acids.
xCT Sodium-independent, high-affinity exchange of anionic amino acids with high specificity for anionic form of cystine and glutamate.
Future directions?
We’d like to learn more.
References
Donsante, A., Johnson, P., Jansen, L. A., & Kaler, S. G. (2010). Somatic mosaicism in Menkes disease suggests choroid plexus-mediated copper transport to the developing brain. American journal of medical genetics. Part A, 152A(10), 2529–2534. PMC free article
Donsante, A., Yi, L., Zerfas, P. M., Brinster, L. R., Sullivan, P., Goldstein, D. S., Prohaska, J., Centeno, J. A., Rushing, E., & Kaler, S. G. (2011). ATP7A gene addition to the choroid plexus results in long-term rescue of the lethal copper transport defect in a Menkes disease mouse model. Molecular therapy : the journal of the American Society of Gene Therapy, 19(12), 2114–2123. PMC free article
Haddad, M. R., Choi, E. Y., Zerfas, P. M., Yi, L., Martinelli, D., Sullivan, P., Goldstein, D. S., Centeno, J. A., Brinster, L. R., Ralle, M., & Kaler, S. G. (2018). Cerebrospinal Fluid-Directed rAAV9-rsATP7A Plus Subcutaneous Copper Histidinate Advance Survival and Outcomes in a Menkes Disease Mouse Model. Molecular therapy. Methods & clinical development, 10, 165–178. PMC free article
Murakami R, Chiba Y, Nishi N, Matsumoto K, Wakamatsu K, Yanase K, Uemura N, Nonaka W, Ueno M. Immunoreactivity of receptor and transporters for lactate located in astrocytes and epithelial cells of choroid plexus of human brain. Neurosci Lett. 2021 Jan 10;741:135479.
Bongarzone, S., Barbon, E., Ferocino, A., Alsulaimani, L., Dunn, J., Kim, J., Sunassee, K., & Gee, A. (2020). Imaging niacin trafficking with positron emission tomography reveals in vivo monocarboxylate transporter distribution. Nuclear medicine and biology, 88-89, 24–33. PMC free article
Spector R. Niacin and niacinamide transport in the central nervous system. In vivo studies. J Neurochem. 1979 Oct;33(4):895-904.
Spector R, Johanson CE. Vitamin transport and homeostasis in mammalian brain: focus on Vitamins B and E. J Neurochem. 2007 Oct;103(2):425-38. Free article
Uchida Y, Goto R, Takeuchi H, Åuczak M, Usui T, Tachikawa M, Terasaki T. (2020) Abundant Expression of OCT2, MATE1, OAT1, OAT3, PEPT2, BCRP, MDR1, and xCT Transporters in Blood-Arachnoid Barrier of Pig and Polarized Localizations at CSF- and Blood-Facing Plasma Membranes. Drug Metab Dispos. 2020 Feb;48(2):135-145. free article
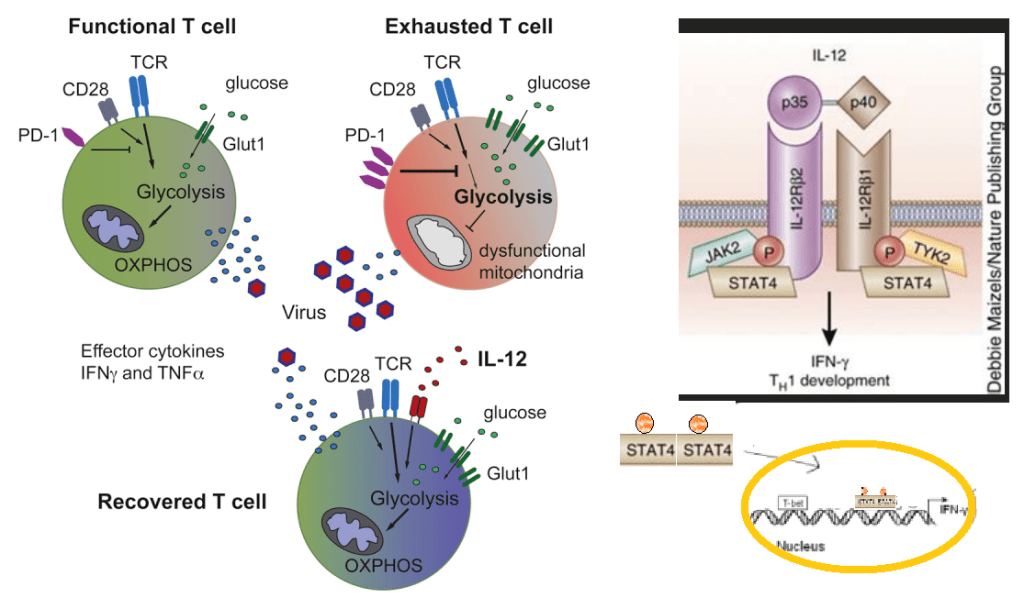
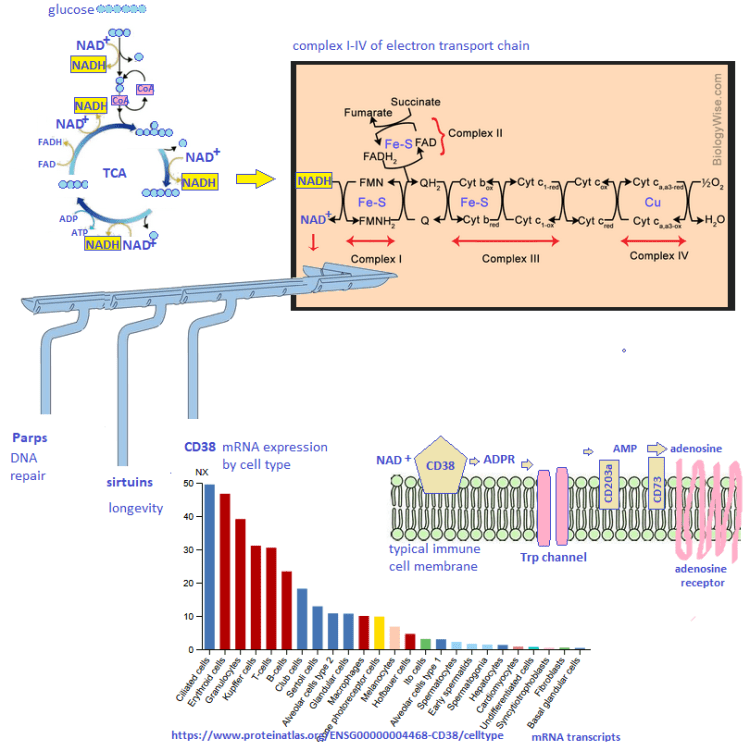
Why do the elderly get sick more than younger folks? Fight Aging has a good lay friendly piece on the mitochondrial aspects of T cell exhaustion. The following paper is cool enough to be published in the prestigious journal Cell. The authors found that hypoxia does something bad to the mitochondria that prevents them from producing cytokines needed to keep chronic virus infections contained. We’ve know for a long time that the real damage to cardiac mitochondria occurs when oxygen is restored. The mitochondria turn into superoxide generating machines. We might be done with the pandemic proper, but older folk are still getting Covid, the flu, and so on. Then there is the chicken pox/ shingles virus.
Schurich A, Pallett LJ, Jajbhay D, Wijngaarden J, Otano I, Gill US, Hansi N, Kennedy PT, Nastouli E, Gilson R, Frezza C, Henson SM, Maini MK. Distinct Metabolic Requirements of Exhausted and Functional Virus-Specific CD8 T Cells in the Same Host. Cell Rep. 2016 Aug 2;16(5):1243-1252. PMC free article
This publication asked the question of why we humans might have some chronic virus infections like cytomegalovirus that are usually well contained and generally harmless. Others, like hepatitis B virus, are far from harmless. The above publication compared CMV and HBV specific T cells from the same patients. The lay reader not particularly intersected in scientific experiments is encouraged to just glance at the images to take in the hard work that goes into understanding out our bodies work.
The graphical abstract of Schurich 2016 has been embellished to demonstrate the role of IL-12 in increasing transcription of interferon gamma.
The patients in this study had both CMV and HBV reactive T cells. Peripheral blood mononuclear cells (PBMC) were isolated from patients positive for HLA-A2+ , one of many variants of the MHC complex of T cell receptors. According to these authors, naïve T cells rely on mitochondrial oxidative phosphorylation of fatty acids. Activated T cells switch to glycolysis as a quicker source of energy even though it is less efficient.
The authors used fluorescent probe labeled peptides from HP,KLHLYSHPI, from Proimmune. As the affinity of isolated, soluble monomeric MHC-peptide complexes for their specific TCR is weak, and the interaction between the MHC-peptide and associated TCR has a half-life of approximately 10 seconds, multimerization is applied to enhance detection sensitivity. The detection sensitivity increases with the amount of MHC monomers complexed onto a multimer. For enhanced detection sensitivity, the Dextramer® technology holds an optimized number of MHC-peptides that enable efficient and sensitive detection of antigen-specific T cells. Read more about the sensitivity of the Dextramer® reagents here.
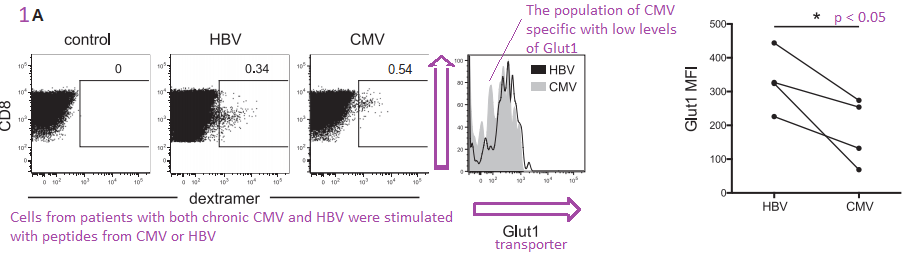
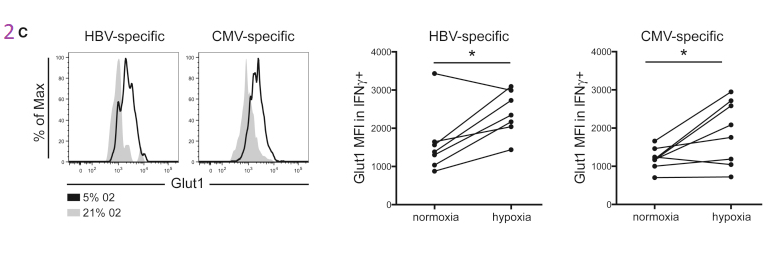
Figure 1. HBV specific CD8+ T Cells express more Glut 1
The authors are building their case that cytotoxic T cells that express the CD8 marker also express more Glut1 because they are more dependent on glycolysis. Virus specific dextramer was used to activate these T cells into cytokine production. The center histogram graph is perhaps the easiest to understand. The CMV specific CD8+ T cell population has a large number of members that just express a few Glut1 transporters on their cell membranes. The implication is that they are more reliant on fatty acid oxidation and OxPhos to meet their energy needs.
Comments on Schurich et al (2016) are in purple.
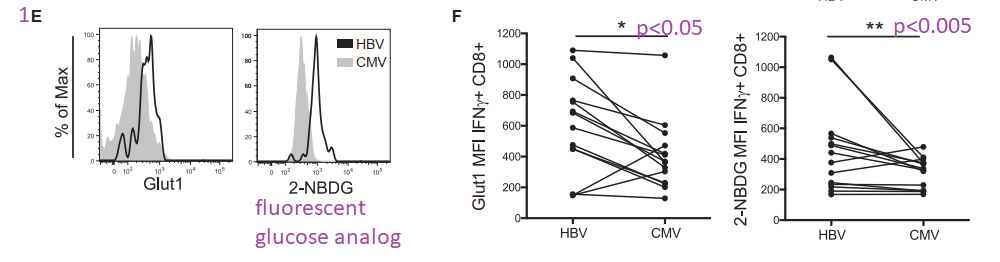
This post is skipping some of the data in Figure 1. Panel 1E asks the question of whether the Glut1 glucose transporters are actually operational. 2-NBDG is glucose with a fluorescent tag NBD. Not only are more Glut1 on the surface of HBV T cells, but they are also operational.
Not the almost non-overlapping of CMV and HBV histograms for 2-NMDG uptake in 1E.
The argument for a copper supplement is that the ATP yield from one molecule is 38 with a copper replete electron transport chain versus 2 ATP from glycolysis alone. The Schurich group has published an excellent review on T cell metabolic requirements in chronic infections. Cholesterol and fatty acid synthesis is needed for T cell proliferation. Fatty acid oxidation was given limited mention. For now we are left with the hypothesis that Glut1 is there because O2 is not available as an electron accept in the electron transport chain. We have to remember that glucose can also be a source of electrons for OxPos ATP production.
Niacin and the role of NADH/NAD+
Our programmed death receptor 1 PD-1 in Long Covid journey began in a previous post. A follow up post examined the role of PD-1 andTim-3 in chronic versus acute LCMV infections. These cell surface receptors shut down transcription of genes for fatty acid metabolism. Sometimes we need to remember niacin, the other two thirds of BioCu1/ Copper One.
This figure is not from Schurich (2016). It is added to illustrate the complexity of NAD+ and ADP/ATP signalling of energy status.
Figure 2. CD8+ T Cells may become hypoxic in the liver.
The next figure compares virus killing cytotoxic T cells from peripheral blood mononuclear populations (PBMC) to intrahepatic liquid (IHL). These cells were stimulated with the HBV dextramer. The low affinity binding (box) is really not that much different. The expression of the Glut1 passive glucose transporter is greater at p<0.05 level of confidence. The authors think that the hepatic environment may be hypoxic. If the liver is hypoxic, could the same situation exist in GI epithelial cells and the SARS-Cov2 virus?
The story continues with PBMC cultured under normoxic and hypoxic conditions. Regardless of the virus specificity, growth in hypoxic conditions increases the expression of Glut1.
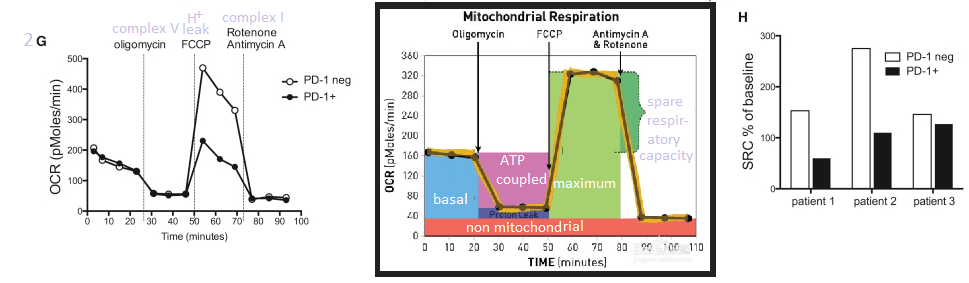
This post makes use of a SeaHorse cartoon illustrating the strategy of gradually adding electron transport chain inhibitors and their use to calculate the spare and maximal respiratory capacity. The spare reserve capacity (SRC) is the maximum O2 consumption minus that which is coupled to ATP production.
Note that the PD-1+ T cells have some sort of electron transport chain defect between complex I and V.
Figure 3 HBV T cells, glycolysis, “immediate effector function”
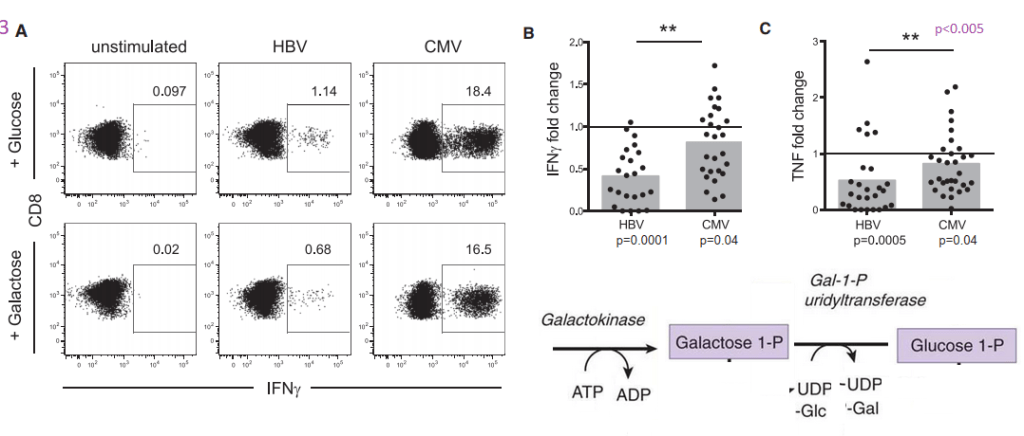
Part of the reasoning behind these experiments was a previous study that Schurich and coauthors cited suggesting that galactose as a sugar source slows down glycolysis. It probably slows down oxidative phosphorylation (ATP production with reduction of oxygen to water. NADH is reduced to NAD+) and the ETC if fatty acid beta oxidation is not a choice.. If sugars are the only source of energy, there is still the additional step required. Glucose to glucose 6-phosphate is the first step in glycolysis. Galactokinase and Gal-1-P uridyltransferase are an additional two steps that slow down glycolysis, but still permit it.
The metabolic diagram in the lower right hand corner was added to the original Schurich figure 3A.
The boxed in areas on the right hand side of the 3A charts are CD8+ T cells that contain a large amount of IFNγ cytokine, presumably in secretory vesicles. Note that T cells that have receptors for CMV have a tendency to express large amounts of IFNγ whether or not the sugar in the growth medium is glucose or galactose. Panel 3B puts numbers to go with the flow plots. The same trend for greater TNF cytokine production is also seen for CMV vs HBV.
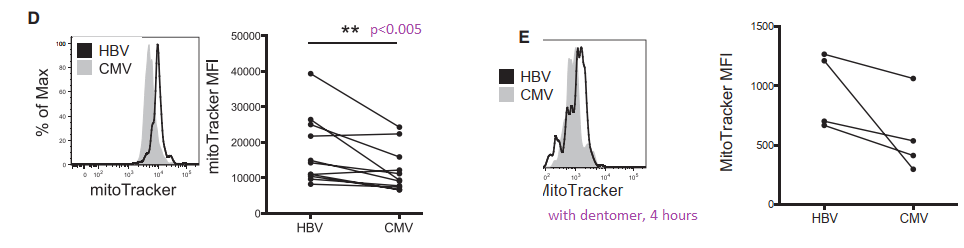
MitoTracker stains healthy and unhealthy mitochondria
The fluorescent dye MitoTracker stains mitochondria regardless of whether or not they have a resting membrane potential. In some pathologies postulated by the authors, mitochondria may become enlarged. The possibility of mitochondrial proliferation was not discussed but could potentially be detected in this assay.
In panel D we see a larger population of HBV specific T cells have more MitoTracker dye uptake. Is this pathology or simply a failed attempt to keep up with increased energy demands. When the T cells are activated with HBV dentomers, the difference is lost.
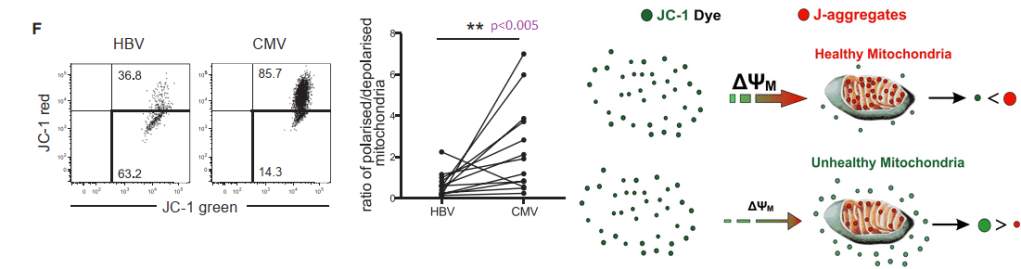
JC-1 measures membrane potential of healthy mitochondria.
The JC-1 mitochondrial membrane potential dye was used to look at functionality.
When the membrane potential is inside negative, JC-1 enters the healthy mitochondria and aggregates in such a way to yield red fluorescence. In HBV virus specific T cells, cells with 50/50 green/red fluorescence predominate, 63.2 %. In CMV specific cells.
Figure 4 IL-12 for exhausted T cells not producing IFNγ?
This post is skipping the work with IL-12. In so many ways Anna Schurich and fellow researches published an interesting study that raises more questions about what is going on in the mitochondria in T cells, or any cells, during hypoxia. Bumping up production of INFγ with IL-12 is interesting. That IL-12 also seems to increase the output of mitochondria is more interesting and leaves us searching for an explanation of why this is.
Why is the liver hypoxic and the many organs that CMV infects not hypoxic?
Is the liver always hypoxic or just when the metabolic load of a meal arrives via the portal vein or when the patient takes a drug that requires metabolism by cytochrome P450?
Could the real mitochondria damage occur when O2 is restored?
Cu(I)NA2 comes to the rescue
Marin W. et al. (2021) Mitochondria as a therapeutic target for cardiac ischemia‑reperfusion injury (Review) Int, J. Mol Med. 47: 485-499, 2021 PMC free article
The review summarizes several decades of research towards the understanding of how cardiac mitochondria become generators of superoxide when ischemia that results from a myocardio infarction is reversed too quickly by reoxygenation. Complex I becomes a generator of super oxide. The Marin review discusses therapeutic interventions to restore the NAD+ that is depleted in the ischemic heart. Nicotinamide riboside (NR) and nicotinamide mononucleotide (NMN) are the natural precursors of NAD+. Copper is only given brief mention in this review as a cofactor in Cu/Zn SOD 1 that removes the superoxide generated by the ischemia reperfusion injured cardiac mitochondria.
Conclusion
The pandemic might be in the rear view mirror for many, but the elderly are still coming down with Covid and many other viral infections that are less of a problem for the young. The lay reader is not expected to understand all of the experiments leading to T cell mitochondria as being the smoking gun for T cell exhaustion in viral infections. We think that the combination of niacin and copper might one day prove to be therapeutic.
What is red light therapy? Why would you or your patient want to take a copper supplement if you are using red light therapy? This post is intended for physicians or for patients to share with their physicians.
What is red light therapy according to Amazon.com?
Red light therapy comes in two wavelengths of LED bulbs: 660 nm and 850 nm, the latter being in the near infrared region of the spectrum. Most of the Amazon treatments are advertised as “FDA cleared.”
stimulate hair growth
wrinkle ablation, skin repair, and overall skin health
muscle and joint pain relief.
Back in 1997 researchers at the Rocheseter General Hospital irradiated isolated rat liver mitochondria fith 660 nm light from an argon dye laser. [1] They found increased activities of NADH:ubiquinone oxidoreductase (ComplexI), ubiquinol: ferricytochrome C oxidoreductase (Complex III) and ferrocytochrome C: oxygen oxidoreductase (Complex IV) (0.6 J/cm2, 1.2 J/cm2, 2.4 J/cm2 and 4.8 J/cm2, P < 0.05). The activities of succinate ubiquinone oxidoreductase (Complex II), ATPase (Complex V), and lactate dehydrogenase were not affected by photo irradiation. [1] Many compounds that transport electrons also absorb visible light. These include heme groups and Fe-S centers in complexes I-IV. While complex IV has two heme groups, only complex IV has copper. Trying to determine if these compounds absorb at 660 and 820 nm is difficult. Complex IV lives in phospholipid membranes rather than in free solution. Red wavelengths of light tend to scatter more.
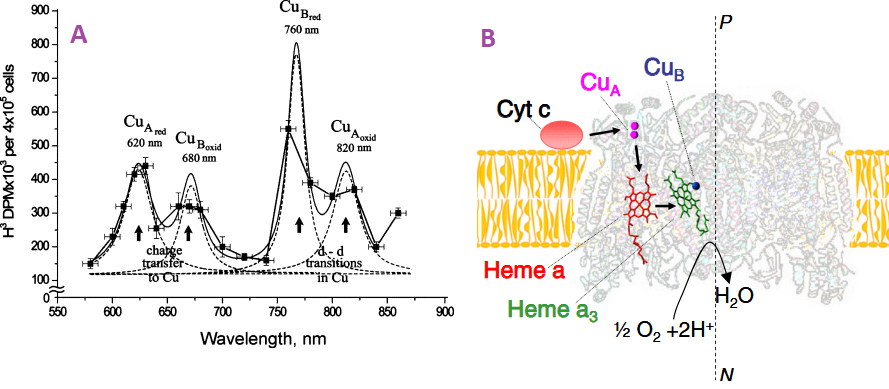
Figure 1. The A. The action spectrum for stimulation of DNA synthesis rate on cellular level. Suggested absorbing chromo phores of the photo acceptor, cytochrome c oxidase, are marked from a review of Tiina Daru [1] Note the Y-axis is not dimensional less absorbance units that many scientists are used to seeing. B. Light absorbing cofactors in complex IV with nitric oxide bound [2].
Figure 1A [2] shows what appears to be an absorption spectrum. It is not. The authors are measuring H3 incorporation into DNA in cells grown in heavy water. H32O. Dr Karu has gone into greater detail as what these “action spectra” are all about. This online publication tells us that Dr Karu made many of these assignments prior to the X-ray structure of cytochrome C oxidase.
Figure 2, Adapted from a Karu online publication. Note the change in absorbance after irradiation with 830nm changes the absorption spectra. The noisy lines are the actual absorbance spectra. The smooth, dashed lines are curve fitting estimations.
820 nm band belongs mainly to oxidized CuA, prior work by Karu, Cu2+-formate, a surrogate for Cu2+ interacting with a carboxyl group, absorbs in this region.
the 760 nm band to reduced CuB, the
680 nm band to oxidized CuB
620 band to reduced CuA, free Cu2+-(his)2 complexes have absorption peaks at 645nm.
400-450 nm band is more likely to be the envelope of a few absorption bands in the range 350-500 nm (i.e., a superposition of several bands).
404-420 nm oxidized heme,
450 nm region, reduced CuB.
Some experiments…
This red light therapy has been FDA approved for wrinkles, wound healing, and hair growth. Would a MitoCopper cream enhance the red light therapy? Would it kick up the mitochondria in those hair follicles into high gear or simply supply copper to lysyl oxidase?These experiments will not answer the big questions but will hopefully tell you whether or not the MitoCopper cream is working for you. If you truly wanted to be scientific, you’d use four treatments: (1) no treatment, (2) just red light, (3) just MitoCopper cream, (4) MitoCopper cream with red light.
Assuming that you do not want to draw squares on your hair thinned scalp with a black Sharpie, find landmarks on your scalp to guide in the application of your copper cream. Leave a similar sized region as a control. Use your red light as per the instructions. Take photos each day before applying the copper cream and using the red light.
If you are using red light therapy to dissipate post pregnancy stretch marks, Pick a particular set of marks to apply CopperOne cream. Take photographs before applying the cream and using the red light.
References
Yu W, Naim JO, McGowan M, Ippolito K, Lanzafame RJ. (1997) Photomodulation of oxidative metabolism and electron chain enzymes in rat liver mitochondria. Photochem Photobiol. 1997 Dec;66(6):866-71
Tiina I Karu TI (2010) Multiple roles of cytochrome c oxidase in mammalian cells under action of red and IR-A radiation. IUBMB Life 2010 Aug;62(8):607-10. free article
Sarti P, Forte E, Mastronicola D, Giuffrè A, Arese M. Cytochrome c oxidase and nitric oxide in action: molecular mechanisms and pathophysiological implications. Biochim Biophys Acta. 2012 Apr;1817(4):610-9. PMC free article
Bhattacharya, M., & Dutta, A. (2019). Computational Modeling of the Photon Transport, Tissue Heating, and Cytochrome C Oxidase Absorption during Transcranial Near-Infrared Stimulation. Brain sciences, 9(8), 179. PMC free article
We have been asked, “Can CopperOne help Amyotrophic lateral sclerosis (ALS) patients given the rumored success of CuATSM clinical trials? This Australian study was accepting patients with familial or sporadic ALS without specifying which proteins (SOD1, TDP-43, or others) are mutated in the familial version when this post was initiated. As of July 25 2023 the results on clincialtials.gov were returned to quality control issues.
Methods: The ALD pilot study was just that. All 12 patient remained on the ALS drug riluzole. Many of the analyses were performed after the patients had died. [1]
CuATSM and riluzole ALS-TDP (n=5) and ALS-SOD1 (n=1)
riluzole only ALS-TDP (n=4) and ALS-SOD1 (n=2)
Results: Our results revealed no significant difference in neuron density or TDP-43 burden in the motor cortex and spinal cord of patients that had received CuATSM compared to patients that had not. In patients that had received CuATSM, p62-immunoreactive astrocytes were observed in the motor cortex and reduced Iba1 density was found in the spinal cord. However, no significant difference in measures of astrocytic activity and SOD1 immuno reactivity was found with CuATSM treatment. [1]
Discussion: CuATSM does not significantly alleviate neuronal pathology or astrogliosis in patients with ALS.
Let’s go back to the original CopperOne customer question, “Can CopperOne help patients with ALS?” We have not clue! We need to first understand what this protein TPD-43 is, why it mis folds, and what may be done to prevent it’s mis folding. We’ll meet a few new copper proteins along the way.
Mutant, misfolded super oxide dismutase 1(SOD1) protein is commonly associated with ALS. An earlier study suggested that sporadic cases were associated with TDP-43 aggregates that tended not to appear in aggregates familial ALS caused by mutations in Cu/Zn SOD1. [2] Misfolded Tar DNA binding protein (TDP-43) is found in many, as well as some familial, cases of ALS. A search of the protein database of crystal structures reveals few structures with single stranded DNA and many aggregates.
The structure of TDP-43
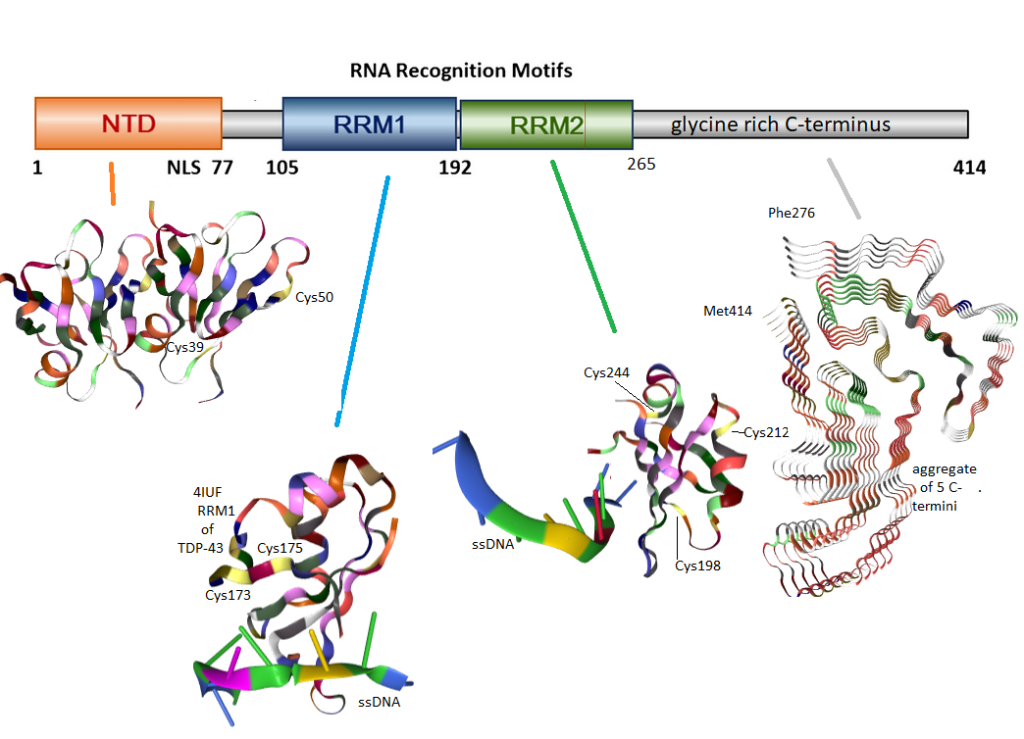
Figure 1 Domains of TDP-43. These images were obtained from rcsb.org. Colors correspond to the type of amino acid. Cysteines are yellow. The N-terminus of TCP-43 has two cysteines which may take the native dimer to a tetramer. The first RNA recognition motif (RRM1) has two cysteines which may form disulfide bonds with each other. RRM2 has three cysteines, two of which have been reported to form aberrant inter-chain cross links. The hydrophobic C-terminus has no disulfides, but can form amyloids as seen in these isolated five fragments.
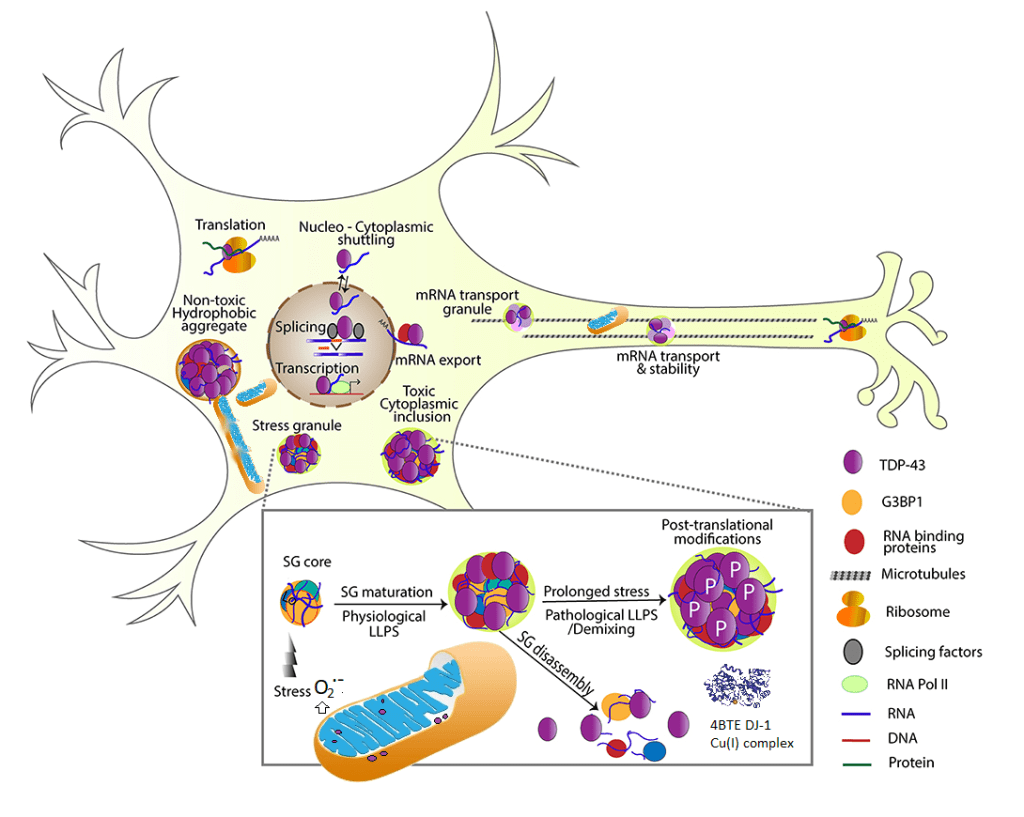
TDP-43 is normally found in the nucleus. A recent review describes its antics in the mitochondria. [2] “TDP-43 proteinopathies†is a term used to describe detergent resistant uniquitinated (figure 2) protein aggregates. As an added bonus, these protein aggregates also tend to be hyper phosphorylated, ubiquitin tags for destruction, and truncated (Figure 1). The accumulation of cytoplasmic inclusion bodies is generally thought of as a toxic gain of function. Formation of these inclusion bodies is accompanied by loss of normal functions that include RNA metabolism,translation of mRNA into protein, stress-induced responses, and mitochondrial function. [3] Studies have demonstrated the association of TDP-43 with mitochondria in motor neuron-like cells. At least a portion of TDP-43 could localize in the inner membrane of mitochondria; full-length and truncated forms of TDPâ€43 could differential reside in the matrix and inter membrane space of mitochondria. [3]
TDP-43 controls mitochondrial trafficking along axons [3], Figure 2
Motor neuron like cells over expressing TDP-43 have reduced mitochondrial complex I activity, mitochondrial transmembrane potential difference, and increased expression of mitochondrial uncoupling protein 2 (UCP2) that functions to decrease the mitochondrial membrane potential.[3]
TDP-43 interacts with mitochondrial RNA
TDP-43 may also facilitate autophagy (self eating) destruction of damaged mitochondria. [2]. The authors mention the Dj-1 protease. As we shall see later, Cu2+ is toxic to proteolytic function.
DJ-1, aka Park7,is a copper cofactor protease localized both in cytoplasm and mitochondria that protects against oxidative stress-induced cell death through the suppression of cytoplasmic TDP-43 aggregation. It was proposed that that the copper enzyme DJ1 may alleviate TDP-43-caused toxicity by degrading both cytoplasmic and mitochondrial TDP-43. [2]
Mitochondrial fission and fusion were discussed in this review [3].
Figure 2 A summary of the many functions of TDP-43. Most of this figure came from [3] with elements of [2] digitally introduced. TFP-43 is mostly a nuclear protein whose interaction with RNA of serves a variety of useful functions [3]. TDP-43 may make its way into the mitochondria where it may disrupt complex 1 as well as disrupt fission and fusion [2]. Ribsomal complexes may also be transported along the axons [3] We suggest that super oxide from dysfunctional mitochondria [2] may release super oxide that is the “stress” trigger for stress granule (SG) core that leads to physiological liquid/liquid phase separation (LLPS).
In a cell culture model Dj-1 co-expression with TDP-43 reduces protein aggregation in response to the mitochondrial toxin and free radical generator paraquat. [4]
TDP-43 Disulfides!
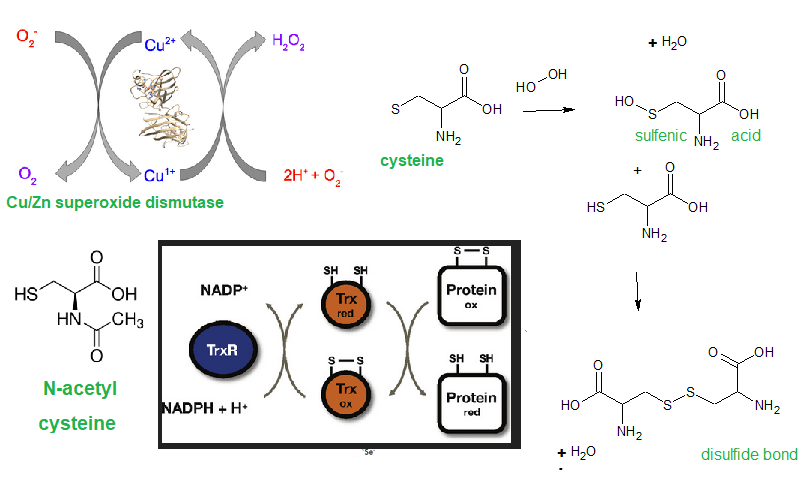
A disulfide is formed when sulfur atoms between two cysteine amino acids form an S-S bond. These disulfide bonds tend to be structural and can be thought of as structural as the buttons and button holes in a dress shirt. Sometimes disulfides form correctly as the new protein comes off the ribosome. In other instances protein disulfide isomerase “unbuttons and rebuttons” disulfides of misfolded proteins in the endoplasmic reticulum. Aberrant disulfides can be formed when sulfur of cysteine is oxidized to cysteine sulfenic acid. This intermediate plays a role in activation of the IL-1β producing inflammasomes.
Figure 3 Cu/Zn SOD 1/3 can be a source of disulfide forming H2O2 as well as detoxifying the much more reactive super oxide. As long as the cell as adequate amounts of catalase, H2O2 will also be detoxified. Thioredoxin (Trx) can use reducing equivalents NADPH + H+ to reduce disulfide bonds.
Let’s take a look at some of the aberrant disulfide bonds of TDP-43 that have been reported in the literature.
Lue71m=,Val72 have been shown to be responsible for non-covalent dimer formation and ability to splice the mRNA for the cystic fibrosis chloride channel CFTR. [5] Two cysteines , Cys39 and Cys50, in this N-terminal domain were shown to be for tetramer formation. [6]
Two forms of oxidative stress, H2O2 and sodium arsenite, have been shown to induce TDP-43 disulfide cross-linking in cultured hippocampus neurons and a cell line expressing TDP-43. Mutants in which Cys198 and Cys244 have been mutated to other amino acids cannot form disulfide bonds. [5]
Cys173 and Cys175 were found to produce intra chain disulfides whereas the others formed inter chain disulfide bonds. [7] Sodium arsentite was used as an indirect means of generating reactive species from the mitochondria.
A mechanism has been proposed by which H2O2 oxidizes one of the two cysteines, Cys198 and Cys244, in the second RNA recognition motif (RRM2). [8]
An extraordinarily through review was published in 2019 by Prasad and coauthors. [9] These authors even covered some publications that suggested TNP-43 aggregation could be protective.
The LOPAC®1280 library, a collection of pharma-developed compounds and approved drugs covering most signaling pathways and major drug target classes was applied at 10 µM concentration to the transfected mouse neuroblastoma Neuro2a (N2a) cells for the TDP-43 self-interaction assay. [10] Auranofin was one compound found to prevent aggregate formation. According to PubChem Auranofin is an inhibitor of thioredoxin reductase, an enzyme thought to give tumor cells resistance to the oxidative stress of their environment.[9] On many levels these results are counter intuitive. Have we not been discussing how inter and intra chain disulfide bond formation can seed aggregation?
The ability of protein disulfide isomerase to prevent aggregation of SOD1 and TDP-43 mutants associated with human ALS adds clarity. [11] Co-expression with functional PDI prevented non-nuclear localization and aggregation in a manner that was dependent on a supply of reduced glutathione.
DJ-1 proteolysis revisited
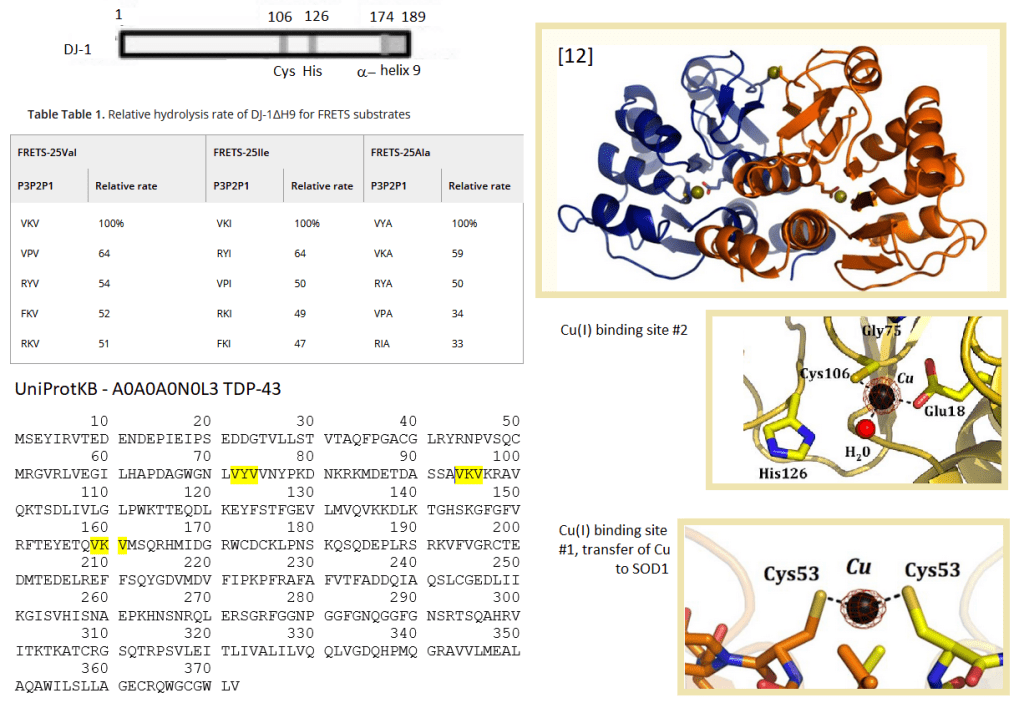
Protease activity of DJâ€1 lacking Câ€terminal αâ€helix (DJâ€1ΔH9) that is cleaved under oxidative stress conditions was stronger than that of fullâ€sized DJâ€1 [12] The most susceptible sequence digested by DJâ€1ΔH9 was valine–lysine–valine-alanine (VKVA).
Figure 4 from reference [11] DJ-1 is a protease auto-inhibited by a C-terminal helix 9. (top) This study uncovered consensus sites for DJ-1 minus the auto-inhibitory C-terminus (middle). The UniProt sequence of human TDP-43 (bottom) reveals that these DJ-1 sites are found the N-terminus of of TDP-43 that include cysteines involved in tetramer-ization. [12] DJ-1 as three Cu sites. (top) The first Cu site may donate Cu to Cu/ZN SOD1. This site is also essential to proteolysis (middle) Cu may be transferred from the first site to the second (bottom)
The optimal conditions of pH 5.5 and 0 mM NaCl. [12]. Nanomolar Cu2+ was inhibitory to DJâ€1’s protease activity. The authors did not comment on monovalent Cu+. Mutation of the first Cu binding site [13] abolished proteolytic activity[12]. DJ-1 loaded with Cu(II) via CuSO4 was shown to photo oxidize to Cu(I). [13] Transfer between Cu(I) from site 1 to site 2 as well as Cu deficient Cu/Zn SOD1 has also been demonstrated. [13] An unanswered question is if Cu(I), but not Cu(II), can be transferred from site 1 to site 2 so as to enable proteolysis.
A thought experiment summary…
Let’s imagine dress shirts in a wash with all sorts of oxidative stress agents that produce cysteine sulfenic acids that act as button hooks facilitating aberrant disulfide bond (button) formation. The pin stripe dress shirt becomes cross-linked to the paisley, which becomes cross-linked to the plain white shirt, which becomes cross-linked to the plaid. .. To make matters worse, the sleeves of the dress shirts become intertwined and form amyloids like the C-termini of TDP-43 in figure 1. The first response would be to add some thioredoxin to unbutton the mess. Partial unbuttoning would only allow the plaid dress shirt to become cross-linked, button to the plain white dress shirt…Thioredoxin inhibitor Auranofin could be added to the mix to keep the dress shirt aggregate from growing larger. PDI, with its thioredoxin like domains, could be added to our dress shirt wash. PDI unbuttons the aberrant buttoning and allows for the right button-button hole matches. Protease DJ-1 reduces our dress shirt aggregation by ripping some of the dress shirts down the back into two pieces. The role of Cu(I) in this process is kind of murky. Cu(I) in DJ-1 [13] can load Cu/Zn SOD with its Cu cofactor so that the aberrant buttoning does not occur in our wash in the first place.
References
Yang Y, Rowe D, McCann H, Shepherd CE, Kril JJ, Kiernan MC, Halliday GM, Tan RH. Treatment with the copper compound CuATSM has no significant effect on motor neuronal pathology in patients with ALS. Neuropathol Appl Neurobiol. 2023 Jun 15:e12919. PubMed
Mackenzie IR, Bigio EH, Ince PG, Geser F, Neumann M, Cairns NJ, Kwong LK, Forman MS, Ravits J, Stewart H, Eisen A, McClusky L, Kretzschmar HA, Monoranu CM, Highley JR, Kirby J, Siddique T, Shaw PJ, Lee VM, Trojanowski JQ. Pathological TDP-43 distinguishes sporadic amyotrophic lateral sclerosis from amyotrophic lateral sclerosis with SOD1 mutations. Ann Neurol. 2007 May;61(5):427-34. Cross Ref
Gao J, Wang L, Yan T, Perry G, Wang X. (2019) TDP-43 proteinopathy and mitochondrial abnormalities in neurodegeneration. Mol Cell Neurosci. 2019 Oct;100:103396. Free PMC article
Loganathan S, Lehmkuhl EM, Eck RJ, Zarnescu DC.(2020) To Be or Not To Be…Toxic-Is RNA Association With TDP-43 Complexes Deleterious or Protective in Neurodegeneration? Front Mol Biosci. 2020 Jan 10;6:154. Free PMC article.
Lei Y, Zhang ZF, Lei RX, Wang S, Zhuang Y, Liu AC, Wu Y, Chen J, Tang JC, Pan MX, Liu R, Liao WJ, Feng YG, Wan Q, Zheng M. DJ-1 Suppresses Cytoplasmic TDP-43 Aggregation in Oxidative Stress-Induced Cell Injury. J Alzheimers Dis. 2018;66(3):1001-1014
Jiang LL, Xue W, Hong JY, Zhang JT, Li MJ, Yu SN, He JH, Hu HY.(2017)The N-terminal dimerization is required for TDP-43 splicing activity. Sci Rep. 2017 Jul 21;7(1):6196. Free PMC article.
Cohen TJ, Hwang AW, Unger T, Trojanowski JQ, Lee VM. EMBO J. (2012) Redox signalling directly regulates TDP-43 via cysteine oxidation and disulphide cross-linking. Embo J.31(5):1241-52. doi: 10.1038/emboj.2011.471. Epub 2011 Dec 23. PMID: 22193716 Free PMC article.
Rabdano SO, Izmailov SA, Luzik DA, Groves A, Podkorytov IS, Skrynnikov NR.(2017) Onset of disorder and protein aggregation due to oxidation-induced intermolecular disulfide bonds: case study of RRM2 domain from TDP-43. Sci Rep. 2017 Sep 11;7(1):11161. Free PMC article.
Prasad A, Bharathi V, Sivalingam V, Girdhar A, Patel BK.(2019) Molecular Mechanisms of TDP-43 Misfolding and Pathology in Amyotrophic Lateral Sclerosis. Front Mol Neurosci. 2019 Feb 14;12:25. doi: 10.3389/fnmol.2019.00025. eCollection 2019. PMID: 30837838 Free PMC article. Review.
Oberstadt M, Stieler J, Simpong DL, Römuß U, Urban N, Schaefer M, Arendt T, Holzer M.(2018)TDP-43 self-interaction is modulated by redox-active compounds Auranofin, Chelerythrine and Riluzole. Sci Rep. 2018 Feb 2;8(1):2248. Free PMC article.
Parakh S, Shadfar S, Perri ER, Ragagnin AMG, Piattoni CV, FogolÃn MB, Yuan KC, Shahheydari H, Don EK, Thomas CJ, Hong Y, Comini MA, Laird AS, Spencer DM, Atkin JD. (2020) The Redox Activity of Protein Disulfide Isomerase Inhibits ALS Phenotypes in Cellular and Zebrafish Models.Science. 2020 May 22;23(5):101097. Free PMC article.
Mitsugi H, Niki T, Takahashi-Niki K, Tanimura K, Yoshizawa-Kumagaye K, Tsunemi M, Iguchi-Ariga SM, Ariga H.(2013) Identification of the recognition sequence and target proteins for DJ-1 protease. FEBS Lett. 2013 Aug 19;587(16):2493-9. doi: 10.1016/j.febslet.2013.06.032. Epub 2013 Jul 2. PMID: 23831022 Free article.
Girotto S, Cendron L, Bisaglia M, Tessari I, Mammi S, Zanotti G, Bubacco L.(2014) DJ-1 is a copper chaperone acting on SOD1 activation. J Biol Chem. 2014 Apr 11;289(15):10887-10899. Free PMC article.